

Table of contents
General morphology - Insertion of the wing - Axillary and basal areas - Venation - The primary veins in the groundplan of Insects - The groundplan of Flies (Longitudinal and crossveins - Primary cells) - Secondary modifications (Costa - Subcosta - Radius - Media - Cubitus - Anals) - Halteres - Bibliography - Web resourcesAbout the morphology and the function performed, the wings are characters of great interest in many orders of insects. The etymology of the name of various orders, on other hands, refers to a recurring feature of the wings. This consideration also applies the Diptera, due some features and the specialization acquired during the evolution.
An apomorphic condition of the Diptera is the transformation of the metathoracic wings into the halteres and the consequent reduction of true wings to a single pair. This feature, present without exception through the entire order, is rarely observed in other insects. The presence of a single pair of wings, however, have distinctive traits which make a fly easily recognizable. Moreover, the etymology of the scientific name of Flies refers to the presence of only two wings.
General morphology

a: tip; an: anal lobe; ax: alula; c: cell; cm: costal margin; fv: false veins; pm: posterior margin; pt: pterostigma; rem: remigium; v: veins.
Author: Giancarlo Dessì
(License: Creative Commons BY-NC-SA)
As in most insects, the wing is derived from a tergopleural expansion which inserts between the tergite and the pleuron and articulates, by axillary sclerites, to special structures derived from the thoracic plates and composed of processes and sclerites. In the Diptera there is a strong differentiation of posterior wings, called halteres, which have got a sharply reduction and profound changes in form and function. So the halteres are transformed into sensory organs accessories of the flight. The forewings mantain the primary function with structural adjustments that bring the ability to flight at the highest levels of efficiency within the class of Insects.
The forewing is divided into three areas, from proximal to distal (McAlpine, 1981):
- the axillary area, which is the proximal end, intimately associated with the tergopleural transition of the thorax; it includes the wing joints;
- the basal stalk, a transitional area which includes the bases of longitudinal veins and their joints with the axillary sclerites;
- the blade, a large area which is the most visible part of the wing. This is the most important for taxonomic purposes, while the other have interest in biological and evolutionary contexts.
The blade is composed of a membranous double layer, bare or more or less pubescent, hyaline or variously pigmented. Inside it there are sclerotized tubules called veins. So there are two structural elements: the venation, composed of the veins, and the membrane. This is composed of a number of more or less large areas called cells, delimited by the veins and the margin. A single cell is closed if completely enclosed by veins, and open if part of the perimeter coincides with the wing margin without veins.

ANP: anterior notal wing process; AxC: axillary cord; N: notum; PN: postnotum; PNP: posterior notal wing process.
Author: Robert Evans Snodgrass (1909)
Modified from the original drawing
(License: Public Domain)
The basic morphological elements of the wing are as follows:
- costal margin: the anterior margin of the wing from the base to the tip; usually it is occupied by the costa, which can reach o go beyond the tip;
- tip: the apex of the wing;
- posterior margin: often mostly membranous, it extends from the tip to the alular incision;
- remigium: the area included between the costal margin, the tip, and the posterior veins derived by the cubitus; it contains the stronger longitudinal veins, as the branches of the radial sector and the media;
- anal lobe: the posterior area, almost entirely membranous, extended between the cubitus and the alular incision;
- pterostigma: not always present, it is a thickening more or less sclerotized and pigmented area near the joint of the anterior branch of radius with the costa;
- disc: the central area of the remigium, often identified by a not basal closed cell called discal cell.
Insertion of the wing
The contents related to anatomy and morphology of the articulation of the wing with the thorax have a great evolutionary and biological importance. The knowledge about these concepts gives interesting approaches about the complexity and the sophistication of a biological function, the flight of insects, which is a high level of biological perfection, and about the studies on the evolution of the articulation system along 300 million years. The comparison between the features of the most primitives insects, as dragonflies and mayflies, and those at the apex of the evolutionary lineages, as flies, bees, beetles and butterflies, gives an idea of the power of nature.

1P, 2P, 3P: first, second and third axillary sclerites; ANP: anterior notal wing process; AxC: axillary cord; Epm: epimeron; Eps: episternum; N: notum; PN: postnotum; PNP: posterior notal wing process; PS: pleural suture; WP: pleural wing process.
Author: Robert Evans Snodgrass (1909)
Modified from the original drawing
(License: Public Domain)
The basic studies about these contents date back to the end of the 19th and the beginning of the 20th century and belong to Snodgrass. In 1909 he publishied a work which published a work which is still one of the most important about the anatomy of the wing insertion. Other studies after Snodgrass (1909) gave improvement of the knowledge, but the basic Snodgrass concepts are still the base of these subject.
At first, the wing is an organ composed of two layers derived from membranous expansions of the tergum and the pleuron. The primary movements which drive the wing during flight (elevation and drop) are given by indirect muscles, the secondary movements, which are important for the functionality of the wing, are given by direct muscles.
The indirect form a strong and complex musculature within the thorax. They are composed of longitudinal, dorsal, and transverse muscles which connect the sclerites of thorax and proximal segments of the legs (tergite, pleuron, sternine, coxae, and trochanters). The combined action of these muscles gives deformations of the thorax which are trasmitted to the wing by the joint of the axillary sclerites and alternately cause the elevation and lowering of the wings.
The direct muscles instead connect the thorax sclerites with those of the axillary region (axillary sclerites, basalare, and subalar sclerite. Their action gives secondary movements of the wing which are important for the folding, during rest, and some functions during the flight, as torsion and flexion. The complexity of the structure of the articulation and the indirect musculature is a primary feature developed throughout the evolution of Neoptera, differentiating them from the primitive Ephemeroptera and Odonata.
In Neoptera, the complex articulation of the wing has a substantial structural uniformity. except specific adaptation of various orders, well described by Snodgrass (1909). This system is composed as follows:
- Wing processes: they are elements of the thoracic sclerites, that support the axillary sclerites by ligaments and joints, and allow the passive transmission of the deformations cause by the indirect muscles. These processes are sclerotized thickening differentiated from the lateral edge of the scutum (anterior notal wing process) and the scutellum (posterior notal wing process) and from the anterodorsal edge of the epimeron (pleural wing process). It is also present a corrugated thickening, the axillary cord, which extends from the posterior border of the notum to the proximal edge of the jugal regione of the wing.
- Thoracic sclerites of the axillary region: they are two small plates, the basalare and the subalar sclerites, derived respectively from the anepisternum and the anepimeron. On them the direct muscles are inserted. The basalare is places in front of the pleural wing process and often it is divided in two smal plates; the anterior basalare (dorsal) and the posterior basalare (ventral). The first one is connected to the scutum, the second one to the anterodorsal of the anepimeron. The subalar sclerite is a thin plate placed in the axillary membrana, behind the pleural wing process.
- Axillary sclerits: they are three (sometimes four) smal sclerites, flattened and with irregular shape, placed in the axillary region of the wing and closely related to the thoracic processes and the basal structures of the wing. They form the real system of the articulation of the wing.
- Sclerites of the basal stalk: they are small plates which connect the wing veins to the axillary sclerites. This plates are distal to the axillary sclerites.
Axillary and basal areas

1: tegula or costal plate; 2: basicosta or humeral plate; 3: subcostal sclerite; 4: first axillary sclerite; 5: second axillary sclerite; 6: third axillary sclerite; 7: stem vein; 8: proximal median plate; 9: distal median plate; 10: anterior notal wing process (mesonotum); 11: pleural wing process (mesopleuron); 12: posterior notal wing process (mesonotum); 13: costagial break; 14: humeral break; 15: lower calypter or squamula thoracica; 16: upper calypter or squamula; 17: alula or axillary lobe; 18: alular incision; 19 anal lobe; A1: first anal; A2: second anal; C: costa; CuA: anterior branch of cubitus; CuP: posterior branch of cubitus; M: posterior media; MA: anterior media; R: radius; Sc: subcosta; a1: anal cell; bc: basal costal cell; bm: basal medial cell; br: basal radial cell; c: costal cell; cup: posterior cubital cell; h: humeral crossvein.
Author: Giancarlo Dessì
(License: Creative Commons BY-NC-SA)
This is a small region including the complex system which connects the wing to the thorax. This area is composed by small sclerites and membranes joined proximally to the pleural sclerites and notal wing processes and distally to the longitudinal veins. They also provide support for the insertion of direct wing muscles. This region is divided into a proximal area, containing the axillary sclerites, and a basal stalk, containing the bases of longitudinal veins. The anatomical structure, rather complicated, is outlined in Figure 4.
The axillary area sensu stricto includes, in anteroposterior and proximal-distal order, these following sclerites:
- Tegula or costal sclerite: the first sclerite at the basal and anterior extreme of the wing. It is distally connected to the basicosta and the costagium.
- Basicosta or humeral sclerite: distal to the tegula and at the front of the costagium is a small elongate plate articulated with the tegula and the base of the costa.
- First axillary sclerite: it has an irregular shape and is placed behind the tegula. It is proximally articulated with the anterior notal wing process of mesothorax, anteriorly with the subcostal sclerite, distally with the second axillary sclerite.
- Second axillary sclerite: it has a transverse vertical devolopment so it connecto both layers of the wing. It is proximally articulated with the first sclerite, anteriorly with the base of stem vein, posteriorly and distally with the third sclerite, and ventrally with the pleural wing process of mesothorax.
- third axillary sclerite: it is proximally articulated with the second sclerite and the posterior notal wing process, distally with the proximal median plate. As the first axillary, it connect to superior layer of the wing.
In some groups, the distal part of the posterior notal wing process is separated from the remaining and becomes a plate of the axillary area forming the fourth axillary sclerite. Within the Diptera this condition occurs onlu in Tabanidae (McAlpine, 1981). When a fourth axillary sclerite is present, this joins to the posterior notal wing process. while the third is joined by the fourth.
The basal stalk is distally close to the axillary area and includes some elongates plates and the basal sections of longitudinal veins. In anteroposterior and proximal-distal order, there are the following elements:
- Costagium: it is the basal section of the first longitudinal vein (C) and it extends from the medial end, joined to the tegula, to the costagial break (if present). The costagium does not connect the axillary sclerites directly, probably to allow the folding of the wing.
- Subcostal sclerite: joined to the base of subcosta, it is included between the tegula, the costagium, the base of stem vein, and finally the first axillary sclerite. Its proximal end is joined to the first axillary so it allows the articulation of the costal region of the wing by merging the subcosta.
- Stem vein: some Authors called it remigium inapropriately. It is the basal section of the radius and is a short and strong vein joined to the first axillary sclerite and distally it connects the radius by a suture-like constriction near the insertion of MA. The articulation with the 2nd axillary sclerite allows the connection of the remigium to the thorax.
- MA or arculus: it is a very short transverse vein confined in the basal stalk. It starts from the M root and reaches the basal section of the radius. This vein could be the vestigial remaining of the ancestral anterior media (McAlpine, 1981).
- Proximal median plate: it is an elongate sclerite oblique to longitudinal axis of the wing closed to the anal region, between the stem vein and the alula. Its proxima end articulates with the 3rd axillary, while its distal end is merged with the distal median plate by a fold.
- Distal median plate: it is distal to the proximal median plate, of which it is the continuation. The distal plate gives the articulation to media, cubitus and anal veins, so the median plates and the 3rd axillary form the system which joins the posterior region of the wing to the thorax.
The main membranous areas of the basal stalk are composed of the basal costal cell (bc), the alula, and the calypteres.
The alula, called also axillary lobe, is a posterior expansion of the membrane, usually well developed in most brachycerous families and reduced or lacking in most of Nematocerous. It is delimited anteriorly by the median sclerites and laterally by two indentations more or less deep: the proximal one separates the alula from the upper calypter, the distal one, called alular incision, from the anal lobe.
The upper calypter, or squamula alaris, is a membranous axillary expansion involved in the movements of the wing and is located between the alula, the third axillary sclerite, and the posterior notal wing process; it is considered homologous of the jugal region of other Neoptera- The lower calypter, or squamula thoracica, is the membranous lobe that connects the posterior margin of the wing to the mesonotum; unlike the upper calypter it is fixed. The calypteres are separated by an indentation of the margin more or less deep. When the wing is at rest, the posterior edge is folded so that the upper calypter covers at least in part the lower. In most Brachycerous, the upper calypter is larger than the lower, but in some groups this ratio is inverted. This condition occurs in most Calyptratae and in some families of lower Brachycerous, such as Tabanidae.
Venation
The venation is a morphological element of basic importance in the taxonomic diagnosis, so the dipterologists pay special attention to the interpretation of the homologies and the resultig terminology. Other fields of application, of the study of the venation, concern the phylogeny, the evolution, and the paleontology. The main problem that recurs in the literature is the controversial interpretation of some homologies, due to a strong reduction of the venation in the Diptera, a trend that occurs in the more advanced orders of insects with membranous wings (Diptera and Hymenoptera): the loss of both longitudinal and crossveins makes it difficult to identify the origin of certain veins and generates incongruity of the terminology adopted in the taxonomic descriptions. Since the late 19th century to today various models of venation scheme are developed; they consist of implementations and adaptations of the original system developed by Comstock & Needham (1898-1899). After the publication of the Manual of Nearctic Diptera (1981) a wide consistency of terminology has emerged, althoug there are still different interpretations by some Authors (Byers, 1989; Saigusa, 2006; Amorim & Rindal, 2007).
The primary veins in the groundplan of Neoptera

A1-3: first, second, third anal; C: costa; CuA: anterior cubitus; CuA1-2: branches of the anterior cubitus; CuP: posterior cubitus; h: humeral; M: media (posterior branch); M1-4: branches of the posterior media; MA: anterior media or arculus or phragma; R: radius; R1: anterior branch of the radius; Rs: radial sector; R2-5: branches of the radial sector; Sc: subcosta; Sc1-2: branches of the subcosta.
Author: Giancarlo Dessì
(License: Creative Commons BY-NC-SA)
Regardless to the importance given to wings since the first attempts about the classification, in the second half of the 19th century a wide literature about this morphologica feature was developed. These studies regard the anatomy and the morphogenesis of the wings. In these works two issues emerged in particular: the first one concerns the morphogenetic origin of the venation, the second one regards the definition of a uniform terminology. These issues were treated specially by Adolph (1879) and Redtenbacher (1886), and later they were reviewed by Comstock and Needham along a decade (Comstock, 1918).
About the morphogenesis, it is believed that the veins are residues of the hemocoel containing tracheae (McAlpine, 1981). During the postembryonic development, at the nymph or pupa stages, the tracheae pervade into the blood lacunae, between the two layers of the wing, and make the traces where the vein is built, by a chitinization of the membrana. This process causes a convex profile of the vein with respect to the plane of the membrana. At a later stage other secondary veins are added. These have a concave profile. Authors usually mark with (+) le convex profile and with (-) the concave. Comstock (1918) found the convex profile as an important feature to interpret the homology in insects with have a reduced venation (p.a. the Flies). After Comstock, the attempts of interpretation of the homologies by dipterists used just this feature during a century, specially in disputes. Adolph (1879) proposed the hypothesis that the longitudinal veins are alternately convex or concave. Comstock and Needham studie in deep this theory and they develop a critical review beginning from the original tracheation of the wing. In summary, all the primary longitudinal veins, except for Sc, have a convex profile, thus they originate from the primitive wing tracheation. The next dichotomous division of certain of these veins originates two branches: the anterior keeps the convex profile, the posterior takes a concave profile.
The question about the terminologic standard was treated by Redtenbacher (1886), who proposed the following names for the six longitudinal veins (from anterior to posterior): Costa, Subcosta. Radius, Media, Cubitus, Anal. Next time, Comstock and Needham defined a nomenclature based on abbreviations and numerical indices according to strict criteria to identify the dichotomous subdivisions. In the Comstock-Needham system, the six longitudinal veins are so named: C, Sc, R, M, Cu, A.
The Comstock e Needham terminology is not in conflict with the names defined by Redtebbacher, since Comstock (1918) suggested the adoption of long names proposed by Redtenbacher as equivalent alternative to abbreviations. Other conventions proposed by Comstock and Needham will be mentioned below.
Each of these primary vein, at least in theory, divides into two branches, the anterior, with convex profile (+), and the posterior one, concave (-). The subcosta is an exception, because it has a concave profile. Each branch arises from a common hemocoelic sinus and may be divided in secondary branches. However this condition occurs only in fossil forms of primitive insects, while in existing forms there is a more or less reduction, specially in higher groups of Neoptera. The structural adataptions that we can encounter are the following:
- atrophy or loss of primary or secondary branches;
- coalescence or anastomosis of two or more branches;
- loss of primitive concavity.
In particular, in Neoptera we find the following conditions:
- The costa is a simple undivided vein.
- The subcosta is usually unbranched, but in primitive forms (Neuropteroidea, Mecoptera) it divides often into two terminal branches referred to as Sc1 and Sc2[1].
- The radius divides into two branches, called R1, unbranched, and radial sector (Rs). The latter have two further dichotomous divisions and gives rise to four branches (R2, R3, R4, and R5).
- The media loses the primary basal stem and seems arising from the base of the cubitus (Figure 4). The anterior media (MA) is strongly reduced and becomes a basal crossvein, called also arculus or phragma, that reaches the base of the radius. The posterior media (MP or usually M) has two dichotomous division into four branches (M1, M2, M3, and M4).
- The cubitus divides into two branches. The anterior (CuA) divides further into CuA1 and CuA2 branches, while the posterior (CuP) remains unbranched.
The groundplan of Flies
In Diptera, at least in the ancestral condition, the venation takes up much of the groundplan of Neoptera, but differs by some features, partly also subject to controversial interpretations.
Longitudinal and crossveins

A1-2: first and second anal; C: costa; CuA: anterior cubitus; CuA1-2: branches of anterior cubitus; CuP: posterior cubitus; h: humeral; M: media (posterior branch); M1-3: branches of posterior media; MA: anterior media; m-cu: medial-cubital; m-m: medial; R: radius; R1: anterior branch of radius; Rs: radial sector; R2-5: branches of radial sector; r-m: radial-medial; Sc: subcosta; sc-r: subcostal-radial.
Author: Giancarlo Dessì
(License: Creative Commons BY-NC-SA)
With regard to the longitudinal veins, in addition to features mentioned for the groundplan of Neoptera, in the Diptera the primitive condition is characterized ad follows.
Costa. It extends along the entire margin, but appears more strong in the costal margin.
Subcosta. In most Diptera, this vein is reduced and reaches C within the proximal third of the costal edge. In some groups it may be broken or coalesced with adjacent veins. In the primitive forms it extends along the entire wing and reaches C near the tip or in the distal third of the costal edge. In Tanyderidae and Tipulomorpha a short vein appears as crossvein between Sc and R1. The homology of this vein is doubtful. Comstock (1918) did not refer to a subcostal-radial crossvein and in a plate about the wing venation of Protoplasa Osten Sacken (Tanyderidae) he pointed this vein as posterior branch of subcosta (Sc2), implicitly admitting the existence even in Diptera. Alexander & Byers (1981b), in the chapter about the Tipulidae followed the Comstock interpretation, referring to this vein as Sc2. But in the general treatment of the same book, McAlpine (1981) cited the existence of a subcostal-radial crossvein (sc-r) and even without considering the homologies, implicitly he excludes the dichotomous division of subcosta in the Diptera. In more recent works, as the Manual of Palaearctic Diptera, there are not specific treatments about this homology, however the thesis of the indivisibility of Sc appears as adopted (Mertz & Haenni, 2000; Dahl & Krzemińska, 1997; Krzemiński & Judd, 1997). It should be noted that the issue of the homology of this vein (Sc2 sensu Comstock or sc-r sensu McAlpine) has not a great importance for taxonomic purposes.
Radius. The base of radius shows a constriction like a suture, approximately near the humeral vein. This constriction separates the stem vein from the rest of the radius. Regarding to the subdivision, it is not entirely certain the keeping of five free branches even in Flies. In most Diptera, the dichotomous subdivision of radius is reduced and only two families of Nematocera (Psychodidae and Tanyderidae) show five apparent free branches. This feature was studied by Alexander in the 20s. About the homologies, where some branches of the radius appear to be coalescent, Alexander believed that the absence of five terminal veins from the radius could be an ancestral condition in the groundplan of Diptera (Alexander, 1929). However, the same Author concluded that there were not sufficient elements to solve the doubt (Alexander, 1929). In fact, in the chapters about Tanyderidae eand Psychodidae in the Manual of Nearctic Diptera, Authors marked the presence of five branche of radius (Alexander, 1981a; Quate & Vockeroth, 1981).
Media. About the structure of the median vein, there are two important features: the reduction of the base, as in the groundplan of Neoptera, and the distal division of the posterior media. Regarding to the latter, in all Diptera, including the most primitive groups, the branching of posterior media and the anterior cubitus originates no more than five veins, instead of six as the ancestral condition of Neoptera (four branches from the media and two from the cubitus). However, in the literature there are two opposite interpretations of the homology of the fourth vein, with important consequences on the nomenclature:

CuA: anterior cubitus; CuA1, CuA1: branches of the anterior cubitus; CuP: posterior cubitus; M1, M2, M3, M4: branches of the posterior media; m-cu: medial-cubitalal.
Author: Giancarlo Dessì
(License: Creative Commons BY-NC-SA)
- the first theory point to the loss of M4 as a free vein. This condition is supported by Comstock (1918), McAlpine (1981), and most Authors in the recent literature. It is explained by the maintenance of the primitive convexity of the last two veins, interpreted as branches of anterior cubitus: if the fourth vein is interpreted as M4, in fact, it should be presumed the inversion of the ancestral concavity of this vein, as resulting from the division of a posterior branch;
- the second hypothesis is based on the presence of M4 as free vein and assumes the invertion of the ancestral concavity. This condition is supported by Tillyard (1926), Hennig (1954, 1973), Colless & McAlpine (1970), and, more recently, by Byers (1989), Saigusa (2006), and Amorim & Rindal (2007). In support of this hypothesis there would be some similarities in the shape of the posterior veins between some families of lower Diptera (Tanyderidae, Tipulidae) and some genera of Mecoptera (Byers, 1989). Therefore, the anterior branch of the cubitus would be undivided or, according to a different thesis, CuA1 would be coalescent with M4.
These different theories are reflected in the nomenclature adopted. Referring to the primitive structure of the venation (see figure 7), there are these following matches:
- the medial-cubital crossvein sensu Comstock (1918) should be the basal section of M4 sensu Tillyard (1926);
- the terminal section of CuA1 sensu Comstock should be the terminal section of M4 sensu Tillyard;
- the basal section of CuA1 sensu Comstock should be the medial-cubital crossvein sensu Tillyard, sometimes referred in literature as tb (Hennig, 1954, 1973);
- CuA2 sensu Comstock should be CuA unbranched sensu Tillyard, sometimes referred as Cu1, or anterior branch of cubitus.
These conflicting theories have different relevance since the twenties. The hypothesis by Tillyard found a wide agreement between the Authors until the eighties, while after the pubblication of the Manual of Nearctic Diptera the interpretation by Comstock has prevailed, due its adoption of McAlpine (1981). However, in recent decades some Authors have followed the Tillyard's model, at least for Nematocera.
Cubitus. About the anterior cubitus, see above. The posterior cubitus is a reduced, weak and incomplete vein; it is also untrachead and close to anterior cubitus.
Anals. The number is reduced to two veins only. The first anal is relatively stout, has a convex shape and usually reaches the margin of the wing. The second anal is relatively weak, incomplete, and concave.
The crossveins present as primitive condition are the following:
- Humeral (h). It is a short vein placed near the base of the wing, between the costa and the subcosta.
- Subcostal-radial (sc-r). Short vein between the subcosta and the anterior branch of the radius (R1). Some Authors interpret it as posterior branch of the subcosta (Sc2).
- Radial (r). It extends between the primary branches of the radius, therefore between R1 and the radial sector or one of its branches (R2+3 or R2). The presence of this vein in the Flies, at least as primitive condition, was supported by Comstock (1918): in the fig. 377, regarding to venation of Tipula, he showed as r a short crossvein between R1 and R2. Otherwise, Alexander suggested that this vein is free section of R2, which after the R2+3 fork it deflects taking a transverse position and finally reaches R1 (Alexander, 1927, 1929, 1981b). It should be pointed that McAlpine (1981) refers this vein as sectoral (r-s), but this statement is inconsistent with the Comstock's terminogy, which defines as sectoral the crossvein extended between the primary branches of the radial sector Comstock (1918).
- Sectoral (s). As noted above, it is a crossvein extended between the primary branches of radial sector, therefore between R2+3 and R4+5 or between R3 and R4 (Comstock, 1918). The keeping of this vein in the Diptera was implicitly suggested by Comstock (1918), in the fig. 358, regarding the venation of Rhynchocephalus Fischer (junior synonym of Nemestrinus Latreille). No sufficient elements there are in literature to support the Comstock's interpretation: the family Nemestrinidae has an unusual venation, within the order, characterized by the presence of vein apparently transverse (referred as "diagonal" in literature) and with uncertain homology. The Authors which treated this family in the Manual of Nearctic Diptera and the Manual of Palaearctic Diptera agree that this veins have originated by elements of radial sector and its branches, r-m crossvein and branches of M and CuA1, while they do not refer to a sectoral sensu Comstock (Teskey, 1981; Richter, 1997).
- Radial-medial (r-m, ta). It is one of the most important crossveins because widely and often mentioned in taxonomic diagnosis. As primary condition, this vein runs between the last branch of the radial sector (R5 or R4+5) and the posterior media. In the past has often been referred as ta (Hennig, 1973).
- Medial (m-m). It is the most distal crossvein, between M2 and M3. The presence of this vein throghout the Diptera is closely related to the interpretation of the homology of CuA1 sensu McAlpine (1981): according to the hypothesis by Comstock (1918), about the subdivision of the anterior cubitus and it coalescence with M4, the m-m crossvein occurs only in primitive groups of Diptera, both Nematocerous and lower Brachycerous; according to hypothesis by Tillyard (1926), about the anterior cubitus unbranched and the keeping of M4, the dm-cu crossvein sensu McAlpine (1981), present in some lower Brachycera and all Cyclorrhapha, could be homologous to m-m of lower Diptera. I do not found references in literature about this personal idea.
- Medial-cubital (strictly m-cu in most Nematocera and Orthorrhapha). In the groundplan based on Comstock (1918) and supported by McAlpine (1981), this vein runs between the basal section of M3 (lower Diptera) and the basal section of CuA1. In the groundplan based on Tillyard (1918) it is identified with the basal section of CuA1 sensu McAlpine before the coalescence with M
4. The Authors that support the hypothesis of the anterior cubitus unbranched refer as medial-cubital crossvein the one interpreted as the basal section of a longitudinal vein by Comstock (1918), derived from the fork of anterior cubitus (Colless & McAlpine, 1970; Byers, 1989; Saigusa, 2006; Amorim & Rindal, 2007).
Primary cells

a1-2: anal; bc: basal costal; bm: basal medial or second basal; br: basal radial or first basal; c: costal; cup: posterior cubital; cua1: anterior cubital; d: disc or discal; m1-3: medial; r1-5: radial; sc: subcostal.
Author: Giancarlo Dessì
(License: Creative Commons BY-NC-SA)
By convention, the nomenclature of the cells is based on the veins that surround them anteriorly, using the lowercase. Referring to the primary venation, in the groundplan there are the following cells:
- Basal costal or first costal (bc): delimited anteriorly by the basal section of costa and distally by the humeral.
- Costal or second costal (c): adjacent and distal to the basal costal, it is delimited by the humeral, the costa and distally by the confluence of the subcosta on the costa.
- Subcostal (sc): delimited anteriorly by the subcosta and posteriorly by R1[2].
- Basal radial or first basal (br): delimited anteriorly by the basal section of radius and the radial sector, posteriorly by the media. Distally it is closed by the radial-medial crossvein.
- Radials (r1, r2, r3, r4, r5): distal to basal radial cell, they are delimited anteriorly by the corresponding branches of the radius. They are usually open cells distally delimited by the margin of wing, but in some groups a radial cell may be closed by coalescences of branches of the radius.
- Basal medial or second basal (bm): in the ancestral condition, according to Comstock (1919) and McAlpine (1981), this cell is delimited anteriorly by the media and posteriorly by the anterior cubitus e and its branch CuA1. Distally it is closed by the medial-cubital crossvein.
- Discal or disc (d): in the groundplan of Diptera, it is the only completely surrounded by other cells, so it is placed more or less in the middle area of the blade. It is delimited anteriorly by M1+2 and posteriorly by M3. Distally it is closed by the medial crossvein (m-m) and proximally by the primary fork of M, so it is adjacent and distal to the basal medial (bm) and proximal to m1 and m2. This cell is reached by the crossveins which close the basal cells: the radial-medial (forward) and the medial-cubital (behind). Depending on the presence and the relative location of M1+2 fork, the discal cell has a pentagonal or hexagonal shape. It recurs in lower Diptera only and it have not to be confused with the discal medial, a secondary cell of upper Diptera, because they are not holomogous.
- Medials (m1, m2, m3): distal to the discal and the basal medial, they are delimited anteriorly by the corrisponding branch of the posterior media and distally by the margin of wing.
- Anterior cubital (cua1): delimited by CuA1 and distally by the margin of wing.
- Posterior cubitale (cup): delimited anteriorly by the posterior cubitus and distally by CuA2 as primitive condition. Some Authors called it as anal cell, but this name is inappropriate because it does not conform to conventions of the Comstock-Needham system. This cell is an important diagnostic character in taxonomy and generally allows to discern between Nematocera and Brachycera. General condition in these groups is the shape of cup cell, open in the first one, closed in the second: the closure of this cell is due to the union of terminal sections of CuA2 and A1, widely recurrent in most of Brachycera. Within the brachycerous Flies, the shape and the extension of this allow to identify some families.
- Anals (a1, a2): they are open cells delimited anteriorly by the corresponding anal vein and distally by the margin of the anal lobe.
Secondary modifications

Veins. A1: anal; C: costa; CuA1-2: branches of anterior cubitus; CuP: posterior cubitus; h: humeral; M1-3: branches of media; m-cu: medial-cubital; m-m: medial; R1: anterior branch of radius; Rs: radial sector; R2-5: branches of radial sector; r-m: radial-medial; Sc: subcosta; sc-r: subcostal-radial.
Cells. bm: basal medial; br: basal radial; cup: posterior cubital; d: discal.
Author: Giancarlo Dessì
(License: Creative Commons BY-NC-SA)
Only a few groups of Nematocera approach at the groundplan as described above. They are the most primitive, as Tipulomorpha and Ptychopteromorpha. Other primitive characters recur in Culicomorpha and Pyschodomorpha. In the rest of Diptera, the venation has evolved so that the specific morphology may deviate considerably from the ancestral condition, sometimes making difficult to interpret the homologies. These adaptations involve substantially the improvement of the functionality of the wings.
In general, the number of divisions of longitudinal veins in the secondary specializations is more reduced than the ancestral condition. In particular, this trend involves the radial sector and the media. A more detailed overview on the most frequent adaptations is summarized in the following sections.
Costa
The extension of the costa to the entire margin remains, as primitive character, in some groups of Nematocera and, rarely, in some Orthorrhapha, while in the generality of Diptera and, in particular, of Brachycera, the costa reaches the end of the radius or the media.
A character particularly important in taxonomy, which occurs mainly in Cyclorrhapha, is the presence of 1-3 breaks of the costa, called costal breaks, corresponding to points of flexibility of the wing. Specific names for these breaks was first introduced by McAlpine (1981):
- costagial break: the most proximal and the least frequent, it is placed between the base of costa and the humeral crossvein. The basal section of the costa, proximal to this break, is called costagium and is usually evident due the presence of short and stout bristles;
- humeral break: rather frequent, it is slightly distal to the humeral crossvein;
- subcostal break: the most frequent, it occurs near the confluence of the subcosta or its projection when this vein is incomplete or fused with R1.
The presence and the number of costal breaks are uniform features at level of single families or higher taxa, so they are useful in taxonomic diagnosis.
Subcosta
Compared to the primitive type, the subcosta of most Diptera has different degrees of simplification and can lose the appearance of a longitudinal vein well formed. Among the changes most frequent there are the following:
- loss of subcostal-radial crossvein sc-r;
- distal section weakly or completely absent;
- distal section coalesced with R1.
Radius

Veins. A1-2: first and second anal; C: costa; CuA1-2: branches of anterior cubitus; h: humeral; M1-3: branches of media; m-cu: medial-cubital; m-m: medial; R1: anterior branch of radius; Rs: radial sector; R2-5: branches of radial sector; r-m: radial-medial; Sc: subcosta.
Cells. bm: basal medial; br: basal radial; cup: posterior cubital; d: discal.
Author: Giancarlo Dessì
(License: Creative Commons BY-NC-SA)
The primitive condition with the division into five branches, with the first from the anterior branch and the other from the radial sector, remains only in Tanyderidae and Psychodidae families. In all other Diptera, the branches of radial sector are reduced to 2-3, except for cases of extreme reduction of the venation. The anterior branch, however, remains present in all groups, although is usually short, specially in Cyclorrhapha.
The nature of the reduction of the branches of the radial sector is uncertain and partly controversial. The examination of the venation within single groups, in fact, induces different interpretations based on the blending of R2 and R3 or, alternatively, on the blending of R4 with R5. For a more detailed analysis of possible homologies, it has to refer to discussion within the single taxonomic groups. These conditions occur in Nematocera and Orthorrhapha, while in upper Brachycera, usually, there is the highest level of simplification, with loss of the second fork of radial sector. So, in these diptera, the radius divides only into three free veins: R1, R2+3 and R4+5.
The sectorial crossvein r appears only in a few groups, but sometimes it is interpreted as blending of R2 on R1. Other special features, recurring in some groups, is the presence of supernumerary crossveins or stumps.
Media
Regardless of different interpretations concerning the loss of M4, as mentioned above, throughout the order it recurs the reduction in number of divisions of posterior media. The extreme condition, typical of Cyclorrhapha, is the presence of only one vein unbranched, usually referred as "media" or sometimes M1 or M1+2.
The probable loss of M3 and the probable coalescence of M1 and M2 have important implications on the morphology of the membrane in the discal area. The presence of a single cell called generically "disc", in the middle of blade, recurs throughout the Diptera, but its genesis differs depending on the systematic group:
- in lower Diptera (Nematocera and Orthorrhapha in part) the presence of M2 and M3 veins, with a medial crossvein between them, involves the closure of a true discal cell. This is the primitive condition as in the groundplan described above;
- in most Cyclorrhaphous and some lower Brachycera, the branches of media and the medial crossvein are lost, and consequently the discal cell is lacking. This condition, usually, is combined with the presence of a secondary medial-cubital crossvein, which closes the cell behind the posterior media. This cell, called medial discal (dm), replaces the primitive true discal.
Under the system proposed by Comstock (1918) and followed by McAlpine (1981), d and dm cells can not be homologous: the true discal is delimited by two primary branches of posterior media (M1+2 anteriorly and M3 posteriorly) and it is distally closed by the medial crossvein (m-m); the medial discal cell, however, is delimited anteriorly by the media and posteriorly by the cubitus (CuA1) and is distally closed by the discal medial-cubital crossvein. Finally, both cells are distal to the basal media cell, but the proximal limit of the true discal is given by the fork of the posterior media, while the proximal limit of the discal medial is given by a crossvein, the basal medial-cubital (bm-cu), which connects the media to the cubitus. The thesis that the primitive discal cell and the secondary discal medial cell are not homologous is further supported by the presence of both cells in some lower Brachycera.
A morphological detail which appears in some groups of Cyclorrhaphous is the partial development or the total disappearance of the basal medial-cubital crossvein, so the basal medial cell is merged with the discal medial making a single closed cell.
Cubitus
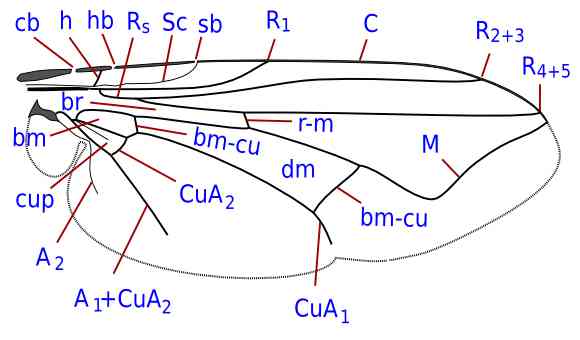
Costal breaks. cb: costagial; hb: humeral; sb: subcostal
Veins. A1-2: first and second anal; bm-cu: basal medial-cubital; C: costa; CuA1-2: branches of anterior cubitus; dm-cu: discal medial-cubital; h: humeral; M: media; R1: anterior branch of radius; Rs: radial sector; R2-5: branches of radial sector; r-m: radial-medial; Sc: subcosta.
Cells. bm: basal medial; br: basal radial; cup: posterior cubital; dm: discal medial.
Author: Giancarlo Dessì
(License: Creative Commons BY-NC-SA)
About the posterior cubitus (CuP) no relevant differences occurs throughout the Diptera compared with the ancestral condition: this vein is weak, untrachead, and incomplete in all Diptera. The only difference that appears is the intensification of the character in upper Diptera, which have the posterior cubitus very weak.
The anterior cubitus has rather a strong secondary differentiation. According to Comstock (1918) and McAlpine (1981), in the primitive flies, this vein gives rise to two branches that reach the posterior margin with a convex trace. This condition recurs in all Nematocerous, except in cases of reduced venation with the loss of one o two branches of cubitus.
In brachycerous Diptera, the vein CuA2 is strongly curved and tends to converge on the first anal. In some Orthorrhaphous, the CuA2 vein is still a free vein and reaches the margin close to anal. In other Orthorrhaphous and the all Cyclorrhaphous, CuA2 bends sharply and converge on A1 until it merges with this vein to form a terminal common section referred as A1+CuA2. This vein may reach or not the edge of wing.
The morphology of CuA2 vein is reflected in the appearance of the posterior cells:
- in Nematocera, the cua1, cup, and a1 cells are well developed and distally limited by the margin of the wing. A general condition of lower Diptera is the shape of cup as open cell;
- in some Orthorrhaphous families (Vermileonidae, Mythicomyiidae, Nemestrinidae, Bombyliidae in part, and Acroceridae in part), the cup cell is open, but tends to closure due the strong convergence of CuA2 and A1 veins;
- in many Orthorrhaphous families, CuA2 ends on the first anal just before the margin, so the cup appears as a closed cell but extended to the edge, which is separated by a short A1+CuA2 vein;
- in all Cyclorrhaphous and some families of Orthorrhaphous, CuA2 is rather short and joins the first anal within the base of wing. The cup appears as a closed and small cell, bordered in the base and usually very shorter than the basal medial cell. The A1+CuA2 vein is relatively long and can reach or not the margin of wing.
The differentiation in shape of this veins and the cup cell is very important as element of taxonomic diagnosis troughout the Brachycerous.
Anals
In the groundplan of Diptera wings, the two anal veins are are complete and free, so reach the margin of the wing delimiting two open cells well defined. This condition remains only in most Tipulomorpha, while in the rest of Diptera a substantial simplification of the venation in the anal lobe recurs.
The first anal is usually weak but well developed and complete. In many groups it is often broken and in the generality of Brachycera is merged with the CuA2 to form a common distal section. When the venation is reduced, the first anal is vestigial or missing.
The second anal, in lower Diptera, is usually weak, poorly developed and incomplete, while in most of Brachycera is vestigial and confined to the base of anal lobe, close to alular incision.
Halteres
The metathoracic wings of Diptera lose their original function and are transformed into the halters, which have function in stabilization of the flight by the perception and the control of the balance of the body. These organs appear as a pedunculated bulb, where we distinguish three parts: scabellum (basal), pedicellum (intermediate) and capitulum (distal and bulbous). They are also called, respctively, base, stem, and knob (McAlpine, 1981).
The scabellum is an expansion articulated on the thorax, rich in sensilla proprioceptors, sensory organs that detect the balance. The pedicellum and the capitulum, due their shapes, adjust the balance of body and stabilize the position during the flight, by vibrations. The pedicellum bears rows of setulae homologous to those of the costa in the forewing.
Due the substantial morphological uniformity, the halteres have limited interest in taxonomy. Any diagnostic elements that are considered are generally limited to the color.
Bibliography
- Adolph, G.E. (1879) Ueber Insektenflügel. Nova Acta der Ksl. Leop.-Carol.-Deutschen Akademie der Naturforscher. Band XLI. Pars II, Nr. 3: 215-291.
- Alexander, C.P. (1927) The interpretation of the radial field of the wing the nematocerous Diptera, with special reference to the Tipulidae. Proceedings of The Linnean Society of New South Wales 52: 42-72.
- Alexander, C.P. (1981) Tanyderidae: 149-151. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera. Volume 1, Work cited.
- Alexander, C.P. & Byers, G.W. (1981) Tipulidae: 152-190. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera. Volume 1, Work cited.
- Amorim, D.S. & Rindal, E. (2007) Phylogeny of the Mycetophiliformia, with proposal of the subfamilies Heterotrichinae, Ohakuneinae, and Chiltrichinae for the Rangomaramidae (Diptera, Bibionomorpha), Work cited.
- Byers, G.W. (1989) Homologies in wing venation of primitive Diptera and Mecoptera, Work cited.
- Cerretti, P. (2010) Terminologia morfologica: 8-11. In I tachinidi della fauna italiana (Diptera Tachinidae), con chiave interattiva dei generi ovest-paleartici. Vol. II. Atlante iconografico, Work cited.
- Colless, D.H. & McAlpine, D.K. (1970) Anatomy of adult: 656. In Diptera (Flies), Work cited.
- Comstock, J.H. & Needham, J.G. (1898-1899) The Wings of Insects, Work cited.
- Comstock, J.H. (1918) The Wings of the Diptera: 347-361. In The Wings of Insects. An Exposition of hte Uniform Terminology of the Wing Veins of Insects and a Discussion of the More General Characteristics of the Wing of the Several Orders of Insects, Work cited.
- Dahl, C. & Krzemińska, E. (1997) 2.9. Family Trichoceridae: 227-237. In Papp, L. & Darvas, B. (eds.) Contributions to a Manual of Palaearctic Diptera. Volume 2. Nematocera and Lower Brachycera, Work cited.
- Hennig, W. (1954) Flügelgeäder und System der Dipteren unter Berücksichtigung der aus dem Mesozoikum berschriebenen Fossilien, Work cited.
- Hennig, W. (1973) Imagines: 141-236. In Diptera (Zweiflüger), Work cited.
- Krzemiński, W. & Judd, D.D. (1997) 2.14. Family Tanyderidae: 281-289. In Papp, L. & Darvas, B. (eds.) Contributions to a Manual of Palaearctic Diptera. Volume 2. Nematocera and Lower Brachycera, Work cited.
- McAlpine, J.F. (1981) Morphology and terminology - Adults: 9-63. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (eds.) Manual of Nearctic Diptera. Volume 1, Work cited.
- Merz, B. & Haenni, J.P. (2000) 1.1. Morphology and terminology of adult Diptera (other than terminalia): 21-51. In Papp, L. & Darvas, B. (eds.) Contributions to a Manual of Palaearctic Diptera. Volume 1. General and Applied Dipterology, Work cited.
- Quate, L.W. & Vockeroth, J.R (1981) Pychodidae: 293-300. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera. Volume 1, Work cited.
- Redtenbacher, J. (1886) Vergleichende Studien über das Flügelgeäder der Insecten. Annalen des K.K. Naturhistorischen Hofmuseums 1 (3): 152-232
- Richter, V.A. (1997) 2.31. Family Nemestrinidae: 460-468. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 2. Nematocera and Lower Brachycera, Work cited.
- Saigusa, T. (2006) Homology of Wing Venation of Diptera, Work cited.
- Servadei, A.; Zangheri, S. & Masutti, L. (1972) Entomologia generale ed applicata, Work cited.
- Snodgrass, R.E. (1909) The thorax of Insects and the articulation of the wings, Work cited.
- Teskey, H.J. (1981) Nemestrinidae: 585-588. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R.; Wood, D.M. (a cura di) Manual of Nearctic Diptera. Volume 1, Work cited.
- Tillyard, R.J. (1926) The Insects of Australia and New Zealand, Work cited.
- Tremblay, E. (1985) Morfologia: 14-24. In Entomologia applicata. Volume Primo: Generalità e mezzi di controllo, Work cited.
- Tremblay, E. (1991) Ordine Diptera (Ditteri): 11-20. In Entomologia applicata. Volume III Parte I, Work cited.
Web resources
- Yeates, D.K.; Hastings, A.; Hamilton, J.; Colless, D.H.; Lambkin, C.L.; Bickel, D.J.; McAlpine, D.K.; Schneider, M.A.; Daniels, G. & Cranston, P.S. Anatomical Atlas of Flies. In CSIRO Entomology. CSIRO, Commonwealth Scientific and Industrial Research Organisation. Last access: 28 May 2019.

Creative Commons BY-NC-SA 3.0 Unported License
(BY: Attribution - NC: Noncommercial - SA: Share Alike).