

Indice della pagina
Morfologia generale - Inserzione dell'ala sul torace - Zona basale-ascellare - Venatura - La venatura ancestrale degli Insetti - La venatura ancestrale dei Ditteri (Nervature longitudinali e trasversali - Cellule primarie) - Modificazioni secondarie (Costa - Subcosta - Radio - Media - Cubito - Anali) - Bilancieri - Bibliografia - Risorse WebIn relazione alla morfologia e alla funzione svolta, le ali sono un carattere di grande interesse per molti ordini di insetti, basti pensare all'etimologia del nome, che spesso richiama una peculiarità ricorrente associata alle ali. Questa considerazione si applica anche ai Ditteri, caratterizzati da alcune peculiarità morfologiche e per il grado di specializzazione acquisito nel corso dell'evoluzione.
Condizione morfologica apomorfica dell'ordine dei Ditteri è la trasformazione delle ali metatoraciche in bilancieri e nella conseguente riduzione del numero di ali funzionanti ad un solo paio. Questa peculiarità, presente senza eccezione alcuna in tutto l'ordine, nel resto degli Insetti si riscontra raramente e, comunque, con caratteristiche distintive tali da rendere indubbiamente identificabile un dittero. Del resto, l'etimologia dello stesso nome scientifico dell'ordine fa riferimento alla presenza di due sole ali.
Morfologia generale

a: apice; an: lobo anale; ax: alula; c: cellula; cm: margine costale; fv: false vene; pm: margine posteriore; pt: pterostigma; rem: regione remigante; v: nervature.
Autore: Giancarlo Dessì
(Licenza: Creative Commons BY-NC-SA)
Come nella generalità degli insetti, l'ala è derivata da un'espansione tergo-pleurale che si inserisce nella zona di connessione fra tergite e pleura e si articola per mezzo di scleriti ascellari a specifiche strutture differenziate dalle due placche toraciche, rappresentate da processi e scleriti. Nei Ditteri, come si è detto, si ha una profonda differenziazione delle ali posteriori, dette bilancieri, che hanno subito una forte riduzione e una modificazione nella forma e nella funzionalità, allo scopo di trasformarsi in organi sensoriali accessori alla funzione del volo. Le ali anteriori mantengono invece la funzione primitiva e hanno subito adattamenti strutturali tale da portare la funzionalità del volo ai più alti livelli di efficienza nell'ambito della classe degli Insetti.
Nell'ala anteriore si distinguono, in senso prossimale-distale, tre zone:
- una zona ascellare, denominata axillary area in McAlpine (1981), costituisce l'estremità prossimale intimamente associata alla zona di transizione tergo-pleurale del torace e comprende le articolazioni dell'ala;
- una zona di transizione basale, denominata basal stalk in McAlpine (1981), comprende i tratti basali delle nervature e le loro articolazioni con gli scleriti ascellari;
- una zona estesa, denominata blade in McAlpine (1981), costituisce la parte più evidente dell'ala. Ai fini della determinazione tassonomica è la regione di maggiore importanza, mentre le altre hanno interesse soprattutto di carattere biologico ed evoluzionistico.
L'area dell'ala è costituita da una doppia lamina membranosa, glabra o più o meno pubescente, ialina o pigmentata, percorsa al suo interno da tubuli sclerificati detti nervature o vene. Strutturalmente vi si distinguono due elementi: la rete di nervature e la membrana. Quest'ultima è morfologicamente composta da areole più o meno estese, dette cellule, delimitate dalle nervature e dal margine. Una cellula si dice chiusa se è interamente delimitata da nervature, aperta se parte del perimetro coincide con il margine non riforzato da una nervatura.

ANP: processo alare anteriore del noto; AxC: cordone ascellare; N: noto; PN: postnoto; PNP: processo alare posteriore del noto.
Autore: Robert Evans Snodgrass (1909)
Riadattato dalla tavola originale
(Licenza: Pubblico Dominio)
Elementi morfologici fondamentali sono i seguenti:
- margine costale: coincide con il margine anteriore dell'ala che si estende dalla base fino all'apice; in genere è rinforzato dalla presenza della costa, che può estendersi fino all'apice o anche oltre;
- apice: è il punto più distale dell'ala;
- margine posteriore: generalmente in buona parte membranoso, si estende dall'apice fino all'incisione alulare;
- regione remigante: è l'area che si estende fra il margine costale, l'apice e le nervature posteriori derivate dalla cubito; è percorsa dalle nervature longitudinali più robuste, che si identificano nelle ramificasioni del settore radiale e della media;
- lobo anale: è una regione posteriore, quasi completamente membranosa, estesa fra la cubito e l'incisione alulare;
- pterostigma: non sempre presente, è un ispessimento più o meno sclerificato e pigmentato, posizionato in corrispondenza della confluenza del ramo anteriore della radio sulla costa;
- disco: è l'area centrale della regione remigante, spesso individuata dalla corrispondenza con una cellula non basale chiusa, detta cellula discale.
Inserzione dell'ala sul torace
Gli aspetti relativi all'anatomia e alla morfologia del sistema di articolazione dell'ala sul torace costituiscono un argomento di grande importanza sia biologica sia evoluzionistica. La conoscenza di questi aspetti offre infatti interessanti spunti sia per la complessità e la sofisticazione di una funzione biologica, il volo degli insetti, che rappresenta un elevato grado di perfezione biologica, sia per lo studio di come i meccanismi di articolazione di questi organi si siano perfezionati nel corso di quasi 300 milioni di anni nel processo evolutivo degli Insetti. Il confronto della struttura dell'articolazione dell'ala tra gli insetti alati più primitivi, come le libellule e le effimere, e quelli all'apice delle linee evolutive, come i ditteri, gli imenotteri, i coleotteri e i lepidotteri, offre un'idea di quanto la natura sia riuscita a superare se stessa.

1P, 2P, 3P: primo, secondo e terzo sclerite ascellare; ANP: processo alare anteriore del noto; AxC: cordone ascellare; Epm: epimero; Eps: episterno; N: noto; PN: postnoto; PNP: processo alare posteriore del noto; PS: sutura pleurale; WP: processo alare pleurale.
Autore: Robert Evans Snodgrass (1909)
Riadattato dalla tavola originale
(Licenza: Pubblico Dominio)
Gli studi fondamentali su questi aspetti risalgono ad un periodo che si estende tra la fine del XIX secolo e l'inizio del XX secolo e si devono all'opera di Snodgrass, che culmina in una pubblicazione del 1909 (Snodgrass, 1909 che, ancora oggi, rappresenta una delle più importanti trattazioni sull'anatomia dell'articolazione dell'ala. Studi successivi, dello stesso Snodgrass o di altri Autori, hanno portato ad un miglioramento delle conoscenze, ma i concetti sviluppati da Snodgrass, oltre un secolo fa, costituiscono ancora oggi la base fondamentale di questa tematica.
In prima analisi, l'ala si può considerare come una struttura composta da due lamine sovrapposte derivate da espansioni membranose del tergite e della pleura. I movimenti che danno impulso ai movimenti primari dell'ala durante il volo (elevazione e abbassamento) sono impressi dai muscoli indiretti, i movimenti accessori, fondamentali per la funzionalità dell'ala, sono invece impressi dai muscoli diretti.
Gli indiretti formano una complessa e robusta muscolatura intrinseca del torace. Sono costituiti da muscoli longitudinali, dorsali, e muscoli trasversali che connettono reciprocamente gli scleriti del torace e dei primi segmenti delle zampe (tergite, pleure, sternite, coxe, trocanteri). L'azione combinata di questi muscoli imprime delle deformazioni al torace che, attraverso le articolazioni degli scleriti ascellari, sono trasmesse alle ali determinandone alternativamente l'elevazione e l'abbassamento.
I muscoli diretti mettono invece in relazione funzionale gli scleriti del torace con quelli della regione ascellare (ascellari, basalare e subalare). La loro azione imprime movimenti accessori dell'ala che sono di grande importanza sia per l'alloggiamento in fase di riposo sia per la funzionalità del volo: sono tali il ripiegamento e la distensione dell'ala, la torsione e la flessione. La complessità dell'articolazione e della muscolatura diretta è un carattere primario sviluppato nel corso dell'evoluzione dei Neoptera, differenziandone sostanzialmente la struttura e la funzionalità rispetto agli Ephemeroptera e gli Odonata, i due ordini più primitivi degli Pterigoti.
Nei Neoptera, il complesso sistema di articolazione dell'ala al torace presenta una sostanziale uniformità strutturale, a meno degli adattamenti specifici dei vari ordini, descritti dettagliatamente da Snodgrass (1909). Questo sistema è così composto:
- Processi alari: sono formazioni proprie degli scleriti del torace, deputate al supporto degli scleriti ascellari, per mezzo di legamenti e articolazioni, e alla trasmissione passiva delle deformazioni impresse dai muscoli indiretti. Si tratta di prominenze ispessite e chitinizzate differenziate dal margine laterale dello scuto (processo alare anteriore del noto) e dello scutello (processo alare posteriore del noto) e dal margine antero-dorsale dell'epimero (processo alare pleurale). A questi si aggiunge un ispessimento corrugato, pari e simmetrico, il cordone ascellare, che si estende posteriormente dal margine del noto per entrare in rapporto con l'estremità prossimale della regione jugale dell'ala.
- Scleriti toracici della regione ascellare: si tratta di due piccoli scleriti, il basalare e il subalare, derivati rispettivamente dall'anepisterno e dall'anepimero, sui quali si inseriscono muscoli diretti. Il basalare è posizionato davanti al processo alare pleurale e spesso è suddiviso in due piccoli scleriti: il basalare anteriore (dorsale), che si collega dorsalmente allo scuto, e il basalare posteriore (ventrale), che si collega al margine antero-dorsale dell'anepimero. Il subalare è un sottile sclerite posizionato nella membrana della zona ascellare, posteriormente rispetto al processo alare pleurale.
- Scleriti ascellari o pterali: sono tre (a volte quattro) piccoli scleriti appiattiti e di forma irregolare, posizionati nella zona ascellare dell'ala e in stretto rapporto con i processi alari del torace e con le strutture basali dell'ala. Rappresentano il sistema vero e proprio con cui l'ala si articola al torace.
- Scleriti della zona basale: sono piccoli scleriti che intermediano l'articolazione delle nervature dell'ala agli scleriti ascellari. Sono disposti in posizione più distale rispetto agli scleriti ascellari.
Zona basale-ascellare

1: tegula o sclerite costale; 2: basicosta o sclerite omerale; 3: sclerite subcostale; 4: primo sclerite ascellare; 5: secondo sclerite ascellare; 6: terzo sclerite ascellare; 7: vena stem; 8: sclerite mediano prossimale; 9: sclerite mediano distale; 10: processo alare anteriore del noto (mesonoto); 11: processo alare pleurale (mesopleura); 12: processo alare posteriore del noto (mesonoto); 13: frattura costagiale; 14: frattura omerale; 15: caliptra inferiore o squama toracica; 16: caliptra superiore o squama alare; 17: alula o lobo ascellare; 18: incisione alulare; 19 lobo anale; A1: prima anale; A2: seconda anale; C: costa; CuA: ramo anteriore della cubito; CuP: ramo posteriore della cubito; M: media posteriore; MA: media anteriore; R: radio; Sc: subcosta; a1: cellula anale; bc: cellula basale costale; bm: cellula basale mediale; br: cellula basale radiale; c: cellula costale; cup: cellula cubitale posteriore; h: vena trasversa omerale.
Autore: Giancarlo Dessì
(Licenza: Creative Commons BY-NC-SA)
È una regione di limitato sviluppo che costituisce il complesso sistema morfoanatomico che raccorda l'ala vera e propria al torace. Questa regione comprende un insieme di piccoli scleriti e di membrane che si articolano da un lato ai processi alari tergopleurali e dall'altro alle nervature longitudinali dell'ala e, infine, offrono il supporto per l'inserzione dei muscoli diretti delle ali. Come detto in precedenza, per convenzione si distingue una zona ascellare, comprendente le articolazioni, e una basale di transizione, comprendente la base delle nervature longitudinali. La struttura anatomica generale, piuttosto complessa, è schematizzata in Figura 4.
La zona ascellare propriamente detta comprende, in ordine antero-posteriore e prossimale-distale, i seguenti scleriti:
- Tegula o sclerite costale: primo sclerite articolare situato all'estremità basale e anteriore dell'ala, in rapporto distale con la basicosta e il costagium.
- Basicosta o sclerite omerale: situato in posizione distale rispetto alla tegula e anteriormente rispetto al costagium, è una placchetta oblunga che si articola alla tegula e alla base della costa.
- Primo sclerite ascellare: di forma irregolare, è posizionato dietro la tegula. Si articola prossimalmente con il processo alare anteriore del noto, anteriormente con lo sclerite subcostale, distalmente con il secondo sclerite ascellare. È in rapporto con la membrana superiore dell'ala.
- Secondo sclerite ascellare: rispetto agli altri ascellari, ha uno sviluppo trasversale in senso dorso-ventrale ed è perciò in rapporto sia con la membrana superiore sia con la membtrana inferiore dell'ala. Si articola prossimalmente con il primo sclerite ascellare, anteriormente con la base della vena stem, posteriormente e distalmente con il terzo sclerite ascellare e, infine, ventralmente con il processo alare pleurale.
- Terzo sclerite ascellare: si articola prossimalmente con il secondo sclerite ascellare e il processo alare posteriore del noto e distalmente con lo sclerite mediano prossimale. Come il primo sclerite, è in rapporto con la membrana superiore dell'ala.
In alcuni gruppi, l'estremità distale del processo alare posteriore del noto si distacca e diventa parte integrante della zona ascellare dell'ala formando il quarto sclerite ascellare. Nei Ditteri, questa condizione ricorre esclusivamente nei Tabanidae (McAlpine, 1981). Quando è presente il quarto sclerite, questo si articola al processo alare posteriore del noto, mentre il terzo ascellare si articola al quarto.
La zona di transizione basale (basal stalk), disposta distalmente rispetto alla zona ascellare, è intimamente connessa a questa e comprende alcuni scleriti, di forma oblunga, e le sezioni basali delle nervature longitudinali. In ordine antero-posteriore e prossimale-distale, gli elementi strutturali di questa regione sono i seguenti:
- Costagium: è il nome specifico con cui si indica la sezione basale della prima nervatura longitudinale (costa) e si estende dall'estremità prossimale, articolata con la tegula fino alla frattura costagiale, quando questa è presente. Il costagium non ha un rapporto articolare diretto con gli scleriti ascellari, condizione probabilmente associata alla funzionalità del ripiegamento dell'ala.
- Sclerite subcostale: fuso con la base della subcosta è compreso fra la tegula, il costagium, la base della vena stem e, infine, il primo sclerite ascellare. Distalmente si articola al primo ascellare e costituisce perciò la connessione articolare della regione costale dell'ala al torace per mezzo della fusione con la subcosta.
- Vena stem: impropriamente chiamata remigium'' da alcuni Autori, costituisce la sezione basale della radio. È una breve e robusta nervatura che si articola prossimalmente al secondo ascellare e si raccorda distalmente con la radio attraverso una strozzatura o una sutura, in corrispondenza dell'intersezione del braccio MA. L'articolazione con il secondo sclerite ascellare conferisce a questa vena un'importante funzione di connessione articolare della regione remigante dell'ala al torace.
- MA o arculus: è una breve vena trasversa confinata nella zona basale, che parte dalla radice della media e si inserisce sulla sezione basale della radio. Questa vena è ritenuta il residuo dell'ancestrale ramo anteriore della media (McAlpine, 1981).
- sclerite mediano prossimale: è una placca oblunga disposta obliquamente rispetto all'asse longitudinale dell'ala, posizionata in prossimità della regione anale, tra la stem e l'alula. La sua estremità prossimale si articola al terzo sclerite ascellare, mentre la sua estremità distale è in rapporto con lo sclerite mediano distale, da cui è morfologicamente distinto per la presenza di una piega di separazione.
- sclerite mediano distale: situato in posizione più distale rispetto allo sclerite prossimale, ne costituisce la continuazione. Su questo sclerite si articolano, in posizioni diverse, le altre nervature longitudinali: la media, la cubito e le anali. L'insieme composto dagli scleriti mediani e dal terzo ascellare forma perciò il sistema attraverso il quale la regione remigante posteriore e la regione anale dell'ala si articolano al torace.
Le principali aree membranose della base sono composte dalla basale costale (bc), dall'alula e dalle caliptre.
L'alula, detta anche lobo ascellare, è un'espansione posteriore della membrana generalmente sviluppata nei Brachiceri e rudimentale o assente nella maggior parte dei Nematoceri. È delimitata anteriormente dagli scleriti mediani e ai lati da due rientranze del margine più o meno profonde: quella prossimale separa l'alula dalla caliptra superiore, quella distale, detta incisione alulare la separa dal lobo anale.
La caliptra superiore, o squama alare, è un'espansione membranosa ascellare coinvolta nei movimenti dell'ala ed è posizionata fra l'alula, il terzo sclerite ascellare e il processo alare posteriore del noto; si ritiene omologa della regione jugale di altri ordini dei Neoptera. La caliptra inferiore, o squama toracica, è il lobo membranoso che raccorda il margine posteriore dell'ala al mesonoto; a differenza della caliptra superiore è fissa. I due lobi sono separati da una incisione del margine più o meno profonda e con il posizionamento dell'ala il margine si ripiega in modo che la caliptra superiore ricopra almeno in parte l'inferiore. Nella generalità dei Brachiceri, la caliptra superiore è più sviluppata della inferiore, ma, in alcuni gruppi, il rapporto dimensionale si inverte. Questa condizione si riscontra nella maggior parte dei Calyptratae e in alcune famiglie di Brachiceri inferiori, come ad esempio i Tabanidae.
Venatura
La venatura dell'ala dei Ditteri è un elemento morfologico di fondamentale importanza nella determinazione tassonomica, perciò si riserva una particolare attenzione all'interpretazione delle omologie e alla conseguente terminologia specifica. Altri ambiti applicativi dello studio della venatura riguardano la filogenesi, l'evoluzionismo e la palentologia. Il problema principale che ricorre in letteratura è l'interpretazione controversa di alcune omologie, a causa di una sostanziale semplificazione della venatura nei Ditteri, fenomeno che ricorre negli ordini più evoluti degli insetti che hanno ali membranose, quali, appunto, i Ditteri e gli Imenotteri: la scomparsa di nervature sia longitudinali sia trasversali rende difficile l'individuazione dell'origine di alcune vene, con conseguente eterogeneità, sotto l'aspetto terminologico, nelle descrizioni ai fini tassonomici. Dalla fine dell'Ottocento ad oggi sono stati sviluppati diversi modelli di riferimento, che consistono in implementazioni e adattamenti dell'originario sistema elaborato da Comstock & Needham (1898-1899). Dopo la pubblicazione del Manual of Nearctic Diptera (1981) si è affermata una sostanziale uniformità della terminologia, per quanto sussistano ancora differenti interpretazioni da parte di alcuni Autori (Byers, 1989; Saigusa, 2006; Amorim & Rindal, 2007).
La venatura ancestrale degli Insetti

A1-3: prima, seconda, terza anale; C: costa; CuA: cubito anteriore; CuA1-2: rami della cubito anteriore; CuP: cubito posteriore; h: omerale; M: media (branca posteriore); M1-4: rami della media posteriore; MA: media anteriore o arculus o fragma; R: radio; R1: ramo anteriore della radio; Rs: settore radiale; R2-5: rami del settore radiale; Sc: subcosta; Sc1-2: rami della subcosta.
Autore: Giancarlo Dessì
(Licenza: Creative Commons BY-NC-SA)
A prescindere dall'importanza data alle ali fin dai primi tentativi di classificazione degli insetti, fu nella seconda metà del XIX secolo che si sviluppò un'ampia letteratura concernente questo carattere morfologico dal punto di vista anatomico e morfogenetico. Nell'ampio panorama dei lavori pubblicati, emersero, in particolare, due problematiche: quella relativa all'origine morfogenetica della venatura alare e quella relativa all'acquisizione di uno standard terminologico. Queste problematiche furono trattate specificamente da Adolph (1879) e Redtenbacher (1886) e, successivamente, rielaborate criticamente da Comstock e Needham nell'arco di un decennio (Comstock, 1918).
Per quanto concerne l'origine morfogenetica, si ritiene che le nervature siano residui del lacunoma contenenti trachee (McAlpine, 1981). Nel corso dello sviluppo postembrionale, nello stadio di ninfa o di pupa, le trachee si insinuano nel lacunoma compreso tra le due lamine che formano la piega dell'ala, delineando la traccia sulla quale viene poi costruita la nervatura, attraverso una chitinizzazione della membrana, che determina la formazione di un profilo convesso della nervatura rispetto al piano della membrana. A queste nervature "primarie" si aggiungono altre nervature, di derivazione secondaria, che hanno invece un profilo concavo. Per convenzione si suole indicare con il simbolo (+) il profilo convesso e con il simbolo (-) il profilo concavo. Sulla base di queste considerazioni, Comstock (1918) individuò nella convessità del profilo un importante elemento su cui basare l'individuazione delle omologie negli insetti con venatura alare ridotta, come ad esempio i Ditteri. È proprio sull'analisi di questo elemento che successivamente si è basato, nell'arco di quasi un secolo, il tentativo di interpretare l'evoluzione della venatura alare nei Ditteri in merito ad alcune incongruenze oggetto di disputa. Adolph (1879) propose una teoria secondo la quale le nervature longitudinali dell'ala sono alternativamente convesse o concave. Comstock e Needham approfondirono in modo critico questa teoria e svilupparono una loro revisione partendo proprio dall'interpretazione dell'originaria tracheazione delle ali. Nella sostanza, tutte le nervature longitudinali primarie, con l'eccezione della subcosta, hanno un profilo convesso e derivano perciò dalla primitiva tracheazione dell'ala. La successiva suddivisione dicotomica di alcune di queste nervature primarie origina due branche, una anteriore, che mantiene il profilo convesso, e una posteriore, che assume un profilo concavo.
Il problema dell'uniformità terminologica fu invece affrontato da Redtenbacher (1886), il quale propose per le sei nervature longitudinali le seguenti denominazioni, in ordine antero-posteriore: Costa, Subcosta. Radio, Media, Cubito, Anale. Successivamente, Comstock e Needham proposero un sistema nomenclaturale basato su abbreviazioni e indicizzazioni numeriche secondo criteri rigorosamente predefiniti per individuare le ramificazioni dicotomiche. Nella fattispecie, le sei nervature primarie, sono denominate, secondo la terminologia di Comstock e Needham, con le seguenti denominazioni abbreviate, in ordine antero-posteriore: C, Sc, R, M, Cu, A.
La terminologia di Comstock e Needham non è in conflitto con quella proposta da Redtenbacher: lo stesso Comstock suggerì l'adozione della denominazione estesa proposta da Redtenbacher come alternativa equivalente ai nomi abbreviati (Comstock, 1918). Nei successivi paragrafi saranno indicate specificamente le convenzioni proposte nel sistema Comstock-Needham.
Ciascuna di queste nervature primarie, almeno in linea teorica, si biforca in due branche, una anteriore, con profilo convesso (+), e una posteriore, con profilo concavo (-). Fa eccezione, come si è detto, la subcosta, che a differenza delle altre nervature primarie ha un profilo concavo. Ciascuna branca trae origine da un seno emocelico comune e a sua volta può ulteriormente dividersi dicotomicamente in rami secondari. Questa condizione, in realtà, si presenta solo in forme fossili di insetti primitivi, mentre in quelle attualmente esistenti si assiste ad una semplificazione più o meno marcata della venatura, in particolare nelle linee più evolute dei Neoptera. Le modifiche strutturali che possono incontrarsi rispetto allo schema ancestrale sono sostanzialmente le seguenti:
- atrofizzazione o scomparsa di branche e rami di primo o secondo ordine;
- fusione o anastomizzazione di branche e rami;
- perdita della primitiva convessità o concavità.
In particolare, nei Neoptera, si riscontrano le seguenti condizioni generali (Comstock, 1918):
- La costa è una nervatura semplice e non subisce divisioni dicotomiche.
- La subcosta è generalmente indivisa, ma nelle forme primitive (Neuropteroidea, Mecoptera) si biforca spesso in due rami terminali indicati con Sc1 e Sc2[1].
- La radio si biforca in due branche, una anteriore (R1), indivisa, una posteriore (Rs o settore radiale). Quest'ultimo subisce due ulteriori suddivisioni dicotomiche originando quattro rami terminali (R2, R3, R4, R5).
- La media perde la sezione basale primaria ed la sua radice è in rapporto con la base della cubito (Figura 4). Subisce una divisione dicotomica originando due branche: la media anteriore (MA) e la media posteriore (MP, a rigore, M sensu Comstock e Needham). Nella generalità degli Insetti, la media anteriore si riduce ad una breve vena trasversale posizionata nella zona basale dell'ala che si unisce alla sezione basale della radio. In letteratura è denominata anche arculus o fragma. La media posteriore, secondo la convenzione terminologica del sistema Comstock-Needham è indicata con il nome M. Questa vena subisce due divisioni dicotomiche originando quattro rami terminali (M1, M2, M3, M4).
- La cubito si divide in due branche secondarie. La cubito anteriore (CuA secondo lo le più recenti convenzioni, Cu1 secondo lo standard originariamente proposto da Comstock e Needham) si divide a sua volta originando i rami CuA1 e CuA2). La cubito posteriore (CuP secondo le convenzioni più recenti, Cu2 secondo lo standard terminologico di Comstock e Needham) resta indivisa.
- Le anali si presentano come semplici vene indipendenti e indivise e individuate con un indice numerico secondo l'ordine antero-posteriore (A1, A2, ...).
La venatura ancestrale dei Ditteri
La venatura dell'ala dei Ditteri, almeno nella condizione primitiva, riprende in buona parte quella dei Neoptera, ma si distingue per alcune peculiarità in parte soggette anche a controverse interpretazioni.
Nervature longitudinali e trasversali

A1-2: prima e seconda anale; C: costa; CuA: cubito anteriore; CuA1-2: rami della cubito anteriore; CuP: cubito posteriore; h: omerale; M: media (branca posteriore); M1-3: rami della media posteriore; MA: media anteriore; m-cu: medio-cubitale; m-m: mediale; R: radio; R1: ramo anteriore della radio; Rs: settore radiale; R2-5: rami del settore radiale; r-m: radio-mediale; Sc: subcosta; sc-r: subcostale-radiale.
Autore: Giancarlo Dessì
(Licenza: Creative Commons BY-NC-SA)
Per quanto concerne le vene longitudinali, in aggiunta agli elementi citati prima per la generalità dei Neoptera, nei Ditteri si riscontrano le condizioni primitive esposte come segue.
Costa. La costa si estende lungo l'intero margine, presentandosi tuttavia più robusta in corrispondenza del margine costale.
Subcosta. Questa nervatura, nella maggior parte dell'ordine, ha uno sviluppo limitato e tende a convergere su C entro il terzo prossimale del margine costale, in diversi gruppi può essere incompleta oppure può fondersi con le vene adiacenti. Nelle forme primitive si estende per buona parte della lunghezza dell'ala, convergendo su C in zona subapicale o, comunque, nel terzo distale del margine costale. Nei Tanyderidae e nei Tipulomorpha è presente una breve vena, all'apparenza trasversa, che si estende da Sc a R1. L'omologia di questa vena è incerta. Comstock (1918) non faceva alcun riferimento all'esistenza di una vena trasversa subcostale-radiale e in una tavola riportante la venatura dell'ala di Protoplasa Osten Sacken (Tanyderidae) indicava questa vena come ramo posteriore della subcosta (Sc2), ammettendone esplicitamente l'esistenza anche nei Ditteri. Alexander & Byers (1981b), nella trattazione dei Tipulidae seguirono l'interpretazione di Comstock, indicando la vena come Sc2. Per contro, nella trattazione generale della stessa opera, McAlpine (1981) cita l'esistenza di una vena trasversale subcostale-radiale e, pur senza entrare nel merito delle omologie, esclude implicitamente la divisione dicotomica della subcosta nella venatura dei Ditteri. In pubblicazioni più recenti, in particolare nel Manual of Palaearctic Diptera, non si riportano trattazioni specifiche di questa omologia, tuttavia si segue implicitamente la tesi dell'indivisibilità della subcosta (Mertz & Haenni, 2000; Dahl & Krzemińska, 1997; Krzemiński & Judd, 1997). Va peraltro precisato che il problema dell'omologia di questa vena (Sc2 sensu Comstock e sc-r sensu McAlpine) è marginale ai fini tassonomici.
Radio. La base della radio presenta una strozzatura, simile ad una sutura, approssimativamente all'altezza della vena omerale, in corrispondenza della zona basale dell'ala. Questa sutura separa distintamente la vena stem dalla radio propriamente detta. Per quanto concerne la ramificazione, vi è incertezza sul mantenimento della suddivisione in cinque rami terminali nei Ditteri. Nella maggior parte dell'ordine la ramificazione dicotomica della radio si semplifica e solo due famiglie di Nematocera (Psychodidae e Tanyderidae) presentano apparentemente cinque rami terminali liberi. Il problema della ramificazione della radio fu studiato in particolare da Alexander negli anni venti. In ragione dell'interpretazione delle omologie negli apparenti casi di fusione di rami della radio, in passato, Alexander sosteneva che l'assenza di cinque terminazioni libere della radio potesse essere una condizione ancestrale da estendersi all'intero ordine dei Ditteri (Alexander, 1929). D'altra parte, lo stesso Autore giunse alla conclusione che non c'erano sufficienti elementi per superare l'incertezza e risolvere definitivamente il problema (Alexander, 1929). In effetti, nella trattazione delle famiglie Tanyderidae e Psychodidae nel Manual of Nearctic Diptera (Alexander, 1981a; Quate & Vockeroth, 1981), gli Autori indicano la presenza di cinque terminazioni libere nella ramificazione della radio.
Media. Sulla struttura della nervatura mediana si evidenziano due caratteri fondamentali: la semplificazione strutturale della base, come descritta nella condizione primitiva dei Neoptera, e della ramificazione terminale della media posteriore. In merito al secondo aspetto, in tutti i Ditteri, comprese le forme più primitive, si originano al massimo cinque vene dalle ramificazioni della media posteriore e della cubito anteriore, contro le sei della condizione primitiva dei Neoptera (quattro rami dalla media e due dalla cubito). In letteratura ricorrono, tuttavia, due differenti e controverse interpretazioni dell'omologia della quarta vena, con importanti implicazioni nella nomenclatura di riferimento:

CuA: cubito anteriore indivisa; CuA1, CuA1: rami della cubito anteriore; CuP: cubito posteriore; M1, M2, M3, M4: rami della media posteriore; m-cu: medio-cubitale.
Autore: Giancarlo Dessì
(Licenza: Creative Commons BY-NC-SA)
- una prima interpretazione considera la scomparsa di M4 come ramo terminale libero. Questa condizione, sostenuta da Comstock (1918), McAlpine (1981) e, nella letteratura degli ultimi decenni, dalla maggior parte degli Autori, si spiega con il mantenimento della primitiva convessità degli ultimi due rami interpretati come suddivisioni della cubito anteriore. Interpretando la quarta vena come M4, infatti, si dovrebbe presuporre l'inversione della concavità primitiva di questo ramo, in quanto derivante dalla suddivisione di una branca posteriore;
- la seconda interpretazione considera invece la presenza di M4 come ramo terminale libero. Questa condizione, sostenuta da Tillyard (1926), Hennig (1954, 1973), Colless & McAlpine (1970) e, più recentemente, da Byers (1989), Saigusa (2006) e Amorim & Rindal (2007), presuppone l'inversione della concavità primitiva di M4. A sostegno di questa ipotesi vi sarebbero alcune affinità nella conformazione della venatura posteriore tra alcune famiglie di Ditteri inferiori (Tanyderidae, Tipulidae) e alcuni generi dei Mecoptera (Byers, 1989). Il ramo anteriore della cubito sarebbe pertanto indiviso oppure, secondo una differente tesi, il ramo CuA1 si sarebbe fuso con M4.
Le due differenti interpretazioni si riflettono sulla nomenclatura adottata. Facendo riferimento alla struttura primitiva della venatura (Figura 7), si hanno le seguenti corrispondenze:
- la vena trasversa m-cu sensu Comstock (1918) corrisponde alla sezione basale di M4 sensu Tillyard (1926);
- la sezione terminale di CuA1 sensu Comstock corrisponde alla sezione terminale di M4 sensu Tillyard;
- la sezione basale di CuA1 sensu Comstock, corrisponde alla vena trasversa m-cu sensu Tillyard (la vena tb sensu Hennig, 1954, 1973);
- la vena CuA2 sensu Comstock corrisponde alla CuA indivisa sensu Tillyard (spesso indicata in letteratura con Cu1, ovvero come branca anteriore della cubito).
Queste interpretazioni controverse hanno avuto differente rilevanza a partire dagli anni venti. L'ipotesi proposta da Tillyard ha infatti trovato un ampio consenso fra i ditterologi fino agli anni settanta, mentre dalla pubblicazione del Manual of Nearctic Diptera ha prevalso l'interpretazione di Comstock, adottata da McAlpine (1981). Va tuttavia ricordato che anche dopo gli anni ottanta sono emerse posizioni a favore dello schema di Tillyard, almeno per i Nematocera.
Cubito. Per quanto riguarda la cubito anteriore, si rimanda a quanto detto poco prima. La cubito posteriore è invece ridotta ad una debole vena incompleta, non percorsa da trachee e strettamente affiancata alla cubito anteriore.
Anali. Nei Ditteri sono ridotte a due sole nervature. La prima anale (A1) è relativamente robusta, ha un profilo convesso e raggiunge il margine. La seconda anale (A2) è invece relativamente debole e incompleta e ha profilo concavo.
Le nervature trasversali presenti come condizione primitiva sono le seguenti:
- Omerale (h). È una breve vena disposta presso la base dell'ala fra la costa e la subcosta.
- Subcostale-radiale (sc-r). Breve vena che si estende dalla subcosta al ramo anteriore della radio (R1). Come detto in precedenza, alcuni Autori la interpretano come ramo posteriore della subcosta (Sc2).
- Radiale trasversa (r). Vena estesa tra le ramificazioni primarie della radio e, quindi, tra R1 e il settore_radiale o una sua ramificazione (R2+3 o R2). La conservazione di questa vena nei Ditteri, almeno come condizione ancestrale, era sostenuta da Comstock (1918), che nel diagramma di fig. 377, riportante la venatura alare nel genere Tipula, indicava con r una breve vena trasversa estesa tra R1 e R2. Alla tesi di Comstock si opponeva quella di Alexander, secondo il quale questa vena doveva interpretarsi come R2, che dopo la biforcazione R2+3 devia assumendo una posizione trasversale per poi fondersi con R1 (Alexander, 1927, 1929, 1981b). Va inoltre precisato che McAlpine (1981) indica l'eventuale vena trasversa estesa tra le ramificazioni primarie della radio con la denominazione settoriale (abbreviato r-s). In realtà, la definizione di McAlpine è incongruente con la terminologia definita da Comstock (1918), che definisce "settoriale" la nervatura trasversa estesa tra ramificazioni primarie del settore radiale.
- Settoriale (s). Come detto poco sopra, è una vena che si estende tra le ramificazioni primarie del settore radiale e, quindi, tra R2+3 e R4+5 oppure tra R3 e R4 (Comstock, 1918). La conservazione di questa vena nei Ditteri è riportata implicitamente da Comstock (1918), che nel diagramma di fig. 358, relativo alla venatura alare del genere Rhynchocephalus Fischer (sinonimo minore di Nemestrinus Latreille). Non ci sono tuttavia sufficienti informazioni, in letteratura, a sostegno dell'interpretazione di Comstock: i Nemestrinidae hanno una venatura singolare, nell'ambito dell'ordine, caratterizzata dalla presenza di nervature apparentemente trasverse (citate come "diagonali" in letteratura) di cui non è facile interpretarne l'omologia. Gli autori che hanno curato la trattazione specifica di questa famiglia nel Manual of Nearctic Diptera e nel Manual of Palaearctic Diptera concordano sull'ipotesi che queste nervature derivino da sezioni del settore radiale e delle sue ramificazioni, della vena r-m, dei rami della media e del ramo CuA1 della cubito anteriore, mentre non si fa alcun riferimento ad una settoriale sensu Comstock (Teskey, 1981; Richter, 1997).
- Radio-mediale (r-m, ta). È una delle nervature trasversali di maggiore importanza per i frequenti riferimenti in sede di determinazione tassonomica. Come condizione primitiva si posiziona fra l'ultimo ramo del settore radiale (R5) e la media posteriore. In passato è stata spesso indicata con con l'abbreviazione ta (Hennig, 1973).
- Mediale (m-m). È la nervatura trasversale più distale, estesa fra i rami M2 e M3 della media posteriore. La ricorrenza di questa vena nell'ambito dell'ordine è strettamente dipendente dall'interpretazione dell'omologia della CuA1 sensu McAlpine (1981): in conformità all'ipotesi di Comstock (1918) sulla biforcazione della cubito anteriore e della sua fusione con M4, la vena m-m ricorre solo in gruppi primitivi di Ditteri, sia tra i Nematoceri sia tra i Brachiceri inferiori; in conformità all'ipotesi di Tillyard (1926) sull'indivisibilità della cubito anteriore e la conservazione del ramo M4 libero, la vena trasversa {{cita|dm-cu}} ricorrente in alcuni Ditteri Ortorrafi e in tutti i Ditteri Ciclorrafi, potrebbe essere omologa alla mediale dei Ditteri inferiori. Su questa interpretazione personale non ho tuttavia trovato riferimenti in letteratura.
- Medio-cubitale (in senso stretto, m-cu nella maggior parte dei Nematocera e degli Orthorrhapha). Nella condizione primitiva teorizzata da Comstock (1918) e sostenuta da McAlpine (1981), questa vena trasversa si posiziona fra la base di M3 e la base di CuA1. In accordo con l'ipotesi di Tillyard (1926), invece, la medio-cubitale si identifica con la sezione basale della vena CuA1 sensu McAlpine prima della presunta fusione con M4. Gli Autori che sostengono la teoria dell'indivisibilità della cubito anteriore, infatti, indicano come medio-cubitale quella che nell'ipotesi di Comstock (1918) è la sezione basale di una nervatura longitudinale derivata dalla biforcazione della cubito anteriore (Colless & McAlpine, 1970; Byers, 1989; Saigusa, 2006; Amorim & Rindal, 2007).
Cellule primarie

a1-2: anali; bc: basale costale; bm: basale mediale o seconda basale; br: basale radiale o prima basale; c: costale; cup: cubitale posteriore; cua1: cubitale anteriore; d: disco o discale; m1-3: mediali; r1-5: radiali; sc: subcostale.
Autore: Giancarlo Dessì
(Licenza: Creative Commons BY-NC-SA)
In conformità ai criteri di nomenclatura definiti nel sistema Comstock-Needhqm, lLa denominazione delle cellule deriva da quella delle nervature che le delimitano anteriormente; per convenzione, la simbologia usa le lettere minuscole. Facendo riferimento alla nervatura primaria, nella condizione ancestrale dei Ditteri ci sono le seguenti cellule:
- Basale costale o prima costale (bc): delimitata anteriormente dalla sezione basale della costa e distalmente dalla omerale.
- Costale o seconda costale (c): adiacente e distale rispetto alla precedente, è delimitata dall'omerale, dalla costa e, distalmente, dalla confluenza della subcosta sulla costa.
- Subcostale (sc): delimitata anteriormente dalla subcosta e posteriormente da R1[2].
- Basale radiale o prima basale (br): è delimitata anteriormente dalla base della radio e dal settore radiale e posteriormente dalla media. Distalmente è chiusa dalla vena r-m.
- Radiali (r1, r2, r3, r4, r5): sono disposte distalmente rispetto alla basale radiale, delimitate anteriormente dalle rispettive ramificazioni della radio. In genere sono distalmente delimitate dal margine dell'ala, ma ricorrono anche casi di cellule radiali chiuse da eventuali coalescenze tra rami della radio.
- Basale mediale o seconda basale (bm): nella condizione primitiva e in accordo con l'interpretazione di Comstock (1918) e McAlpine (1981) è delimitata anteriormente dalla base della media e posteriormente dalla cubito anteriore e dal ramo CuA1. Distalmente è chiusa dalla vena m-cu.
- Discale o disco (d): nella condizione primitiva è l'unica cellula chiusa interamente circondata da altre cellule e, quindi, posizionata più o meno al centro della regione remigante. È delimitata anteriormente dal ramo M1+2 e posteriormente dal ramo M3. È chiusa all'estremità distale dalla vena trasversa m-m e all'estremità prossimale dalla biforcazione primaria della media posteriore. Da questa cellula si estendono le nervature trasversali r-m dal lato anteriore e m-cu dal lato posteriore; in funzione della presenza e della posizione relativa della biforcazione M1+2, ha un profilo pentagonale o esagonale. È presente solo nei Ditteri inferiori (Nematoceri e parte dei Brachiceri Ortorrafi) e non va confusa con la cellula discale mediale (dm), che si forma invece secondariamente nei Ditteri superiori.
- Mediali (m1, m2, m3): sono posizionate distalmente rispetto alla discale e alla basale mediale e sono delimitate anteriormente dalla rispettiva ramificazione della media e distalmente dal margine dell'ala.
- Cubitale anteriore (cua1): è una cellula aperta delimitata anteriormente dalla vena CuA1 e distalmente dal margine dell'ala.
- Cubitale posteriore (cup): è delimitata anteriormente dalla cubito posteriore e distalmente, nei Ditteri inferiori, dalla vena CuA2 e dal margine dell'ala. In letteratura è stata indicata da alcuni Autori come cellula anale, ma questa denominazione è inappropriata in quanto non conforme alla convenzione di nomenclatura del sistema Comstock-Needham. Rappresenta un importante elemento morfologico di determinazione tassonomica e permette, in generale, la differenziazione fra i Nematocera e i Brachycera. Condizione generale di questi due gruppi è infatti la conformazione della cellula cup; nei Nematoceri è una cellula aperta in quanto la vena CuA2 raggiunge il margine dell'ala; nlla generalità dei Brachiceri è invece una cellula chiusa a seguito della coalescenza delle sezioni terminali di CuA2 e A1. Nell'ambito dei Brachiceri, la conformazione e l'estensione di questa cellula può essere utile per una facile individuazione di alcune famiglie.
- Anali (a1, a2): sono cellule aperte delimitate anteriormente dalle rispettive nervature anali e distalmente dal margine del lobo anale.
Modificazioni secondarie

Vene. A1: anale; C: costa; CuA1-2: rami della cubito anteriore; CuP: cubito posteriore; h: omerale; M1-3: rami della media; m-cu: medio-cubitale; m-m: mediale; R1: ramo anteriore della radio; Rs: settore radiale; R2-5: rami del settore radiale; r-m: radio-mediale; Sc: subcosta; sc-r: subcostale-radiale.
Cellule. bm: basale mediale; br: basale radiale; cup: cubitale posteriore; d: discale.
Autore: Giancarlo Dessì
(Licenza: Creative Commons BY-NC-SA)
Allo schema della condizione ancestrale illustrato in precedenza si avvicinano solo pochi gruppi di Nematoceri, che fanno capo ai taxa più primitivi, come i Tipulomorpha e gli Ptychopteromorpha. Altri caratteri evidentemente primitivi ricorrono in parte nei Culicomorpha e negli Psychodomorpha. In tutti gli altri ditteri si assiste ad evoluzioni della nervatura che fanno discostare anche notevolmente la struttura specifica da quella primitiva, talvolta rendendo difficile l'identificazione delle omologie. Questi adattamenti implicano nella sostanza un miglioramento della funzionalità delle ali.
In generale, nelle specializzazioni secondarie si assiste ad una semplificazione più o meno marcata, con la riduzione del numero di ramificazioni delle nervature longitudinali, rispetto al disegno ancestrale. Questa tendenza interessa in modo particolare il settore radiale e la media. Un quadro più dettagliato relativo alle modificazioni secondarie più ricorrenti è riassunto nelle sezioni successive.
Costa
L'estensione della costa all'intero margine resta, come carattere primitivo, in alcuni gruppi di Nematocera e, in misura minore, in alcuni Orthorrhapha, mentre nella generalità dei Ditteri e, in modo particolare, nei Brachiceri, la costa si interrompe in corrispondenza della confluenza di un ramo della radio o della media, in zona apicale o subapicale.
Un carattere di particolare importanza tassonomica, che ricorre soprattutto nei Cyclorrhapha, è la presenza di 1-3 interruzioni della costa, dette fratture costali, corrispondenti a punti di maggiore flessibilità dell'ala. La denominazione specifica per queste fratture è stata introdotta per la prima volta da McAlpine (1981):
- frattura costagiale: è la più prossimale e la meno frequente, disposta fra la base della costa e la confluenza della omerale. La sezione della costa prossimale rispetto a questa frattura è detta costagium e si evidenzia, in genere, per la presenza di robuste e brevi setole;
- frattura omerale: abbastanza frequente, è disposta in posizione leggermente distale rispetto alla confluenza della omerale;
- frattura subcostale: è quella con la maggiore frequenza ed è posizionata in corrispondenza della confluenza della subcosta sulla costa oppure, quando la subcosta è incompleta o fusa con R1, in corrispondenza del punto in cui confluisce la sua proiezione.
La presenza e il numero di fratture costali sono un carattere stabilmente uniforme nell'ambito delle varie famiglie o di taxa di rango superiore, perciò rappresentano un utile elemento di determinazione diagnostica.
Subcosta
Rispetto al tipo primitivo, la subcosta può subire vari gradi di semplificazione fino a perdere quasi del tutto il carattere di vena longitudinale ben marcata. Fra le modificazioni più frequenti che può subire ricorrono le seguenti:
- scomparsa della vena trasversale sc-r;
- evanescenza della sezione terminale fino alla completa scomparsa dell'apice, con trasformazione in una vena incompleta;
- coalescenza della sezione distale con R1.
Radio

Vene. A1-2: prima e seconda anale; C: costa; CuA1-2: rami della cubito anteriore; h: omerale; M1-3: rami della media; m-cu: medio-cubitale; m-m: mediale; R1: ramo anteriore della radio; Rs: settore radiale; R2-5: rami del settore radiale; r-m: radio-mediale; Sc: subcosta.
Cellule. bm: basale mediale; br: basale radiale; cup: cubitale posteriore; d: discale.
Autore: Giancarlo Dessì
(Licenza: Creative Commons BY-NC-SA)
La condizione primitiva della suddivisione in cinque rami, di cui uno anteriore e quattro posteriori derivati dalla suddivisione dicotomica del settore radiale, si mantiene solo nelle famiglie Tanyderidae e Psychodidae. In tutti gli altri Ditteri, il numero di ramificazioni del settore radiale si riduce generalmente a 2-3, fatta eccezione per i casi di estrema riduzione della venatura. Il ramo anteriore, invece, resta presente in tutti i gruppi, anche se spesso, in particolare nei Cyclorrhapha, subisce una marcata riduzione in lunghezza.
La natura della riduzione della ramificazione del settore radiale è incerta e talvolta controversa. L'esame della venatura nei singoli gruppi, infatti, porta a differenti interpretazioni che in sostanza ribadiscono, da un lato, la fusione dei rami R2 e R3, da un altro, la fusione di R4 e R5. Analisi più dettagliate dell'interpretazione delle omologie devono essere rimandate alla trattazione dei singoli gruppi sistematici. Tali condizioni si verificano nei Nematoceri e negli Ortorrafi, mentre nei Brachiceri superiori, in generale, si assiste al più alto grado di riduzione, con la scomparsa della seconda ramificazione dicotomica del settore radiale. Nella generalità di questi Ditteri, la radio si suddivide pertanto in tre rami liberi: R1, R2+3 e R4+5.
La vena trasversale r compare in pochi gruppi, ma è spesso interpretata come confluenza di R2 su R1. Altri caratteri specifici ricorrenti in alcuni gruppi consistono nella presenza di vene trasversali sovrannumerarie o di rami ciechi.
Media
A prescindere dalla divergenza d'interpretazione relativa alla "scomparsa" della vena M4, di cui si è detto in precedenza, nell'ambito dell'ordine si assiste alla riduzione del numero di ramificazioni della media posteriore fino alla presenza, come condizione generale dei Cyclorrhapha, di una sola vena indivisa indicata semplicemente come "media".
La probabile scomparsa del ramo M3 e la probabile fusione dei rami M1 e M2 hanno importanti implicazioni sulla morfologia della membrana nell'area discale dell'ala. In molti Ditteri, è ricorrente la presenza di una cellula centrale, genericamente chiamata "disco", la cui origine, tuttavia, differisce secondo il gruppo sistematico:
- nei Ditteri inferiori (Nematoceri e parte degli Ortorrafi) la presenza di tre rami liberi della media, combinata con la presenza di una vena trasversale mediale, comporta la chiusura della cellula discale propriamente detta. È, questa, la condizione primitiva contemplata nello schema ancestrale descritto in precedenza;
- nella generalità dei Ciclorrafi e in alcuni Ortorrafi, la scomparsa della vena mediale e delle ramificazioni della media comporta, come conseguenza, la scomparsa della cellula discale. Questa condizione, in genere, si accompagna alla comparsa di una vena medio-cubitale secondaria che chiude la cellula posizionata sotto la media. Questa cellula, detta discale mediale (dm), sostituisce la discale primitiva.
Nell'ambito dello schema di Comstock (1918), fatto proprio da McAlpine (1981), le cellule d e dm non possono ritenersi omologhe: la discale propriamente detta è infatti delimitata dalle due ramificazioni primarie della media posteriore (M1+2 anteriormente e M3 posteriormente) ed è chiusa distalmente dalla mediale m-m; la discale mediale, invece, è delimitata dalla media anteriormente e da un ramo cubito anteriore posteriormente (CuA1) ed è chiusa distalmente dalla medio-cubitale discale. Infine, entrambe sono disposte in posizione distale rispetto alla basale mediale, tuttavia la delimitazione prossimale della discale propriamente detta è originata dalla biforcazione primaria della media posteriore, mentre la delimitazione prossimale della discale mediale è originata da una nervatura trasversale, la medio-cubitale basale, che connette la media alla cubito. La tesi che le due cellule non siano omologhe è supportata peraltro dalla loro contemporanea presenza in alcuni gruppi dei Brachiceri inferiori.
Un particolare morfologico che ricorre in alcuni gruppi di Ciclorrafi è lo sviluppo parziale o la completa scomparsa della medio-cubitale basale. Conseguenza di questo adattamento è la fusione della cellula basale con la discale mediale in un'unica cellula chiusa.
Cubito
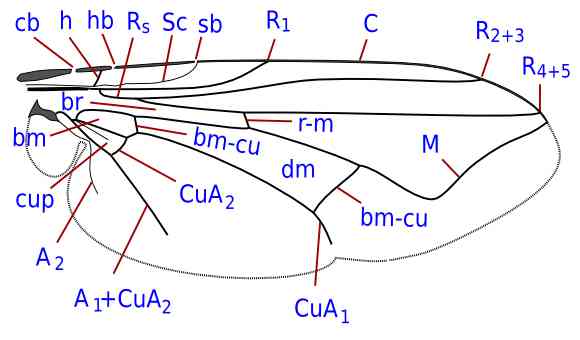
Fratture costali. cb: costagiale; hb: omerale; sb: subcostale
Vene. A1-2: prima e seconda anale; bm-cu: medio-cubitale basale; C: costa; CuA1-2: rami della cubito anteriore; dm-cu: medio-cubitale discale; h: omerale; M: media; R1: ramo anteriore della radio; Rs: settore radiale; R2-5: rami del settore radiale; r-m: radio-mediale; Sc: subcosta.
Cellule. bm: basale mediale; br: basale radiale; cup: cubitale posteriore; dm: discale mediale.
Autore: Giancarlo Dessì
(Licenza: Creative Commons BY-NC-SA)
Per quanto concerne la cubito posteriore (CuP) non si rilevano nell'ordine marcate differenziazioni rispetto alla condizione primitiva: questa vena si presenta debole, priva di trachee e incompleta in tutti i Ditteri. L'unico elemento di differenziazione consiste nell'accentuazione del carattere nei Ditteri superiori, fino a rendere quasi evanescente la cubito posteriore.
La morfologia della cubito anteriore presenta invece una marcata differenziazione secondaria. In accordo con l'interpretazione di Comstock (1918) e McAlpine (1981), nel tipo primitivo, questa nervatura si biforca originando due rami completi che raggiungono il margine con un tracciato convesso rispetto al margine costale. Tale condizione si verifica in generale in tutti i Nematoceri, eccetto i gruppi in cui si ha una semplificazione della venatura, con conseguente scomparsa di uno o più rami della cubito.
Nei Brachiceri, il ramo CuA2 subisce una deviazione più o meno marcata, accentuando la convessità del tracciato, e tende a convergere sulla prima anale. In alcuni Brachiceri Ortorrafi, la vena CuA2 mantiene ancora la primitiva individualità e raggiunge il margine dell'ala, pur assumendo un percorso convergente con quello dell'anale. Nella generalità dei Brachiceri Ciclorrafi e nella maggior parte degli Ortorrafi, invece, la vena CuA2 subisce una forte curvatura e converge sull'anale fino a fondersi con essa per formare una vena terminale A1+CuA2, che può raggiungere o meno il margine dell'ala.
Il comportamento della vena CuA2 si riflette sulla morfologia delle cellule posteriori:
- nei Nematoceri, le cellule cua1, cup e a1 sono tutte ben sviluppate e delimitate distalmente dal margine posteriore dell'ala. Condizione generale dei Ditteri inferiori è la conformazione della cup come cellula aperta;
- in alcune famiglie di Ortorrafi (es. Vermileonidae, Mythicomyiidae, Nemestrinidae, parte dei Bombyliidae e degli Acroceridae), la cellula cup è ancora aperta, ma, a causa della forte convergenza di CuA2 e A1, tende a chiudersi;
- in molte famiglie di Ortorrafi, la CuA2 confluisce sulla prima anale poco prima di raggiungere il margine. La cup si configura perciò come una cellula chiusa ma estesa fino al margine, da cui resta separata dal breve tratto della vena A1+CuA2;
- nella generalità dei Ciclorrafi e in alcune famiglie di Ortorrafi, la CuA2 è piuttosto ridotta in lunghezza e confluisce sulla prima anale nella parte basale dell'ala. La cup si configura perciò come una cellula chiusa e di limitato sviluppo, confinata alla base dell'ala e generalmente molto meno estesa della cellula basale mediale. La vena A1+CuA2 è relativamente lunga e può raggiungere o meno il margine dell'ala.
La differenziazione nella conformazione di queste vene e della cellula cup assume una rilevanza tale da ritenersi un importante elemento di determinazione tassonomica nell'ambito della generalità dei Brachiceri.
Anali
Nella condizione primitiva, le due vene anali sono complete e raggiungono autonomamente il margine dell'ala, delimitando due cellule aperte ma ben delineate. Tale condizione si riscontra solo nella generalità dei Tipulomorpha, mentre nel resto dell'ordine ricorre la sostanziale e ulteriore riduzione della venatura del lobo anale.
La prima anale si presenta in genere come vena poco robusta ma ben conformata e completa. In molti gruppi è spesso incompleta e nella maggior parte dei Brachiceri si fonde, come si è detto, in un tratto terminale comune con la CuA2. In gruppi con venatura ridotta, la prima anale si presenta rudimentale oppure scompare del tutto.
La seconda anale, nei Ditteri inferiori, è in genere debole, poco sviluppata e incompleta, mentre nella maggior parte dei Brachiceri si presenta come una breve vena rudimentale confinata alla base del lobo anale e adiacente all'incisione alulare.
Bilancieri
Le ali metatoraciche dei Ditteri perdono la loro primitiva funzione e sono trasformate in bilancieri, la cui funzione è quella di stabilizzare il volo con la percezione dell'equilibrio e il bilanciamento del corpo. Questi organi hanno la forma di un bulbo peduncolato, in cui si distinguono tre parti: una basale, detta scabellum, una peduncolare intermedia, detta pedicellum, e una terminale, bulbosa, detta capitulum.
Lo scabellum è un'espansione articolata al metatorace ricca di sensilli propriocettori, organi sensoriali preposti alla percezione dell'equilibrio. Il pedicellum e il capitulum, in virtù della loro forma, hanno la funzione di bilanciare il corpo e stabilizzarne la posizione durante il volo per mezzo di movimenti di vibrazione. Il pedicellum è rivestito da file di setole che sarebbero omologhe a quelle inserite sulla costa nelle ali anteriori.
Per la sostanziale uniformità morfologica, i bilancieri hanno scarso interesse dal punto di vista tassonomico. Gli eventuali elementi diagnostici presi in considerazione si limitano in genere alla pigmentazione.
Bibliografia
- Adolph, G.E. (1879) Ueber Insektenflügel. Nova Acta der Ksl. Leop.-Carol.-Deutschen Akademie der Naturforscher. Band XLI. Pars II, Nr. 3: 215-291.
- Alexander, C.P. (1927) The interpretation of the radial field of the wing the nematocerous Diptera, with special reference to the Tipulidae. Proceedings of The Linnean Society of New South Wales 52: 42-72.
- Alexander, C.P. (1929). A comparison of the systems of nomenclature that have been applied to the radial field of the wing in the Diptera. Proceedings of the Fourth International Congress of Entomology (Ithaca, 1928) 2: 700-707.
- Alexander, C.P. (1981) Tanyderidae: 149-151. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera. Volume 1, Op. cit.
- Alexander, C.P. & Byers, G.W. (1981) Tipulidae: 152-190. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera. Volume 1, Op. cit.
- Amorim, D.S. & Rindal, E. (2007) Phylogeny of the Mycetophiliformia, with proposal of the subfamilies Heterotrichinae, Ohakuneinae, and Chiltrichinae for the Rangomaramidae (Diptera, Bibionomorpha), Op. cit.
- Byers, G.W. (1989) Homologies in wing venation of primitive Diptera and Mecoptera, Op. cit.
- Cerretti, P. (2010) Terminologia morfologica: 8-11. In I tachinidi della fauna italiana (Diptera Tachinidae), con chiave interattiva dei generi ovest-paleartici. Vol. II. Atlante iconografico, Op. cit.
- Colless, D.H. & McAlpine, D.K. (1970) Anatomy of adult: 656. In Diptera (Flies), Op. cit.
- Comstock, J.H. & Needham, J.G. (1898-1899) The Wings of Insects, Op. cit.
- Comstock, J.H. (1918) The Wings of the Diptera: 347-361. In The Wings of Insects. An Exposition of hte Uniform Terminology of the Wing Veins of Insects and a Discussion of the More General Characteristics of the Wing of the Several Orders of Insects, Op. cit.
- Dahl, C. & Krzemińska, E. (1997) 2.9. Family Trichoceridae: 227-237. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 2. Nematocera and Lower Brachycera, Op. cit.
- Hennig, W. (1954) Flügelgeäder und System der Dipteren unter Berücksichtigung der aus dem Mesozoikum berschriebenen Fossilien, Op. cit.
- Hennig, W. (1973) Imagines: 141-236. In Diptera (Zweiflüger), Op. cit.
- Krzemiński, W. & Judd, D.D. (1997) 2.14. Family Tanyderidae: 281-289. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 2. Nematocera and Lower Brachycera, Op. cit.
- McAlpine, J.F. (1981) Morphology and terminology - Adults: 9-63. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera. Volume 1, Op. cit.
- Merz, B. & Haenni, J.P. (2000) 1.1. Morphology and terminology of adult Diptera (other than terminalia): 21-51. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 1. General and Applied Dipterology, Op. cit.
- Quate, L.W. & Vockeroth, J.R (1981) Pychodidae: 293-300. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera. Volume 1, Op. cit.
- Redtenbacher, J. (1886) Vergleichende Studien über das Flügelgeäder der Insecten. Annalen des K.K. Naturhistorischen Hofmuseums 1 (3): 152-232
- Richter, V.A. (1997) 2.31. Family Nemestrinidae: 460-468. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 2. Nematocera and Lower Brachycera, Op. cit.
- Saigusa, T. (2006) Homology of Wing Venation of Diptera, Op. cit.
- Servadei, A.; Zangheri, S. & Masutti, L. (1972) Entomologia generale ed applicata, Op. cit.
- Snodgrass, R.E. (1909) The thorax of Insects and the articulation of the wings, Op. cit.
- Teskey, H.J. (1981) Nemestrinidae: 585-588. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R.; Wood, D.M. (a cura di) Manual of Nearctic Diptera. Volume 1, Op. cit.
- Tillyard, R.J. (1926) The Insects of Australia and New Zealand, Op. cit.
- Tremblay, E. (1985) Morfologia: 14-24. In Entomologia applicata. Volume Primo: Generalità e mezzi di controllo, Op. cit.
- Tremblay, E. (1991) Ordine Diptera (Ditteri): 11-20. In Entomologia applicata. Volume III Parte I, Op. cit.
Risorse Web
- Yeates, D.K.; Hastings, A.; Hamilton, J.; Colless, D.H.; Lambkin, C.L.; Bickel, D.J.; McAlpine, D.K.; Schneider, M.A.; Daniels, G. & Cranston, P.S. Anatomical Atlas of Flies. In CSIRO Entomology. CSIRO, Commonwealth Scientific and Industrial Research Organisation. Ultimo accesso: 28 maggio 2019.

Licenza Creative Commons BY-NC-SA
(BY: Attribuzione - NC: Non commerciale - SA: Condividi allo stesso modo).