

Indice della pagina
Caratteri della cuticola (Sclerificazione - Pigmentazione) - Escrescenze - Bibliografia - Risorse WebIn questa pagina prendiamo in esame i caratteri principali associati al tegumento delle larve, ovvero il grado di sclerificazione e la pigmentazione della cuticola, le caratteristiche degli annessi cuticolari di consistenza non carnosa, rappresentati da tricomi di vario sviluppo e aspetto come peli, setole, spine, ecc. Si rimanda invece alla pagina successiva la descrizione dei processi molli, che, pur apparendo come appendici del corpo, sono in realtà estroflessioni carnose della cuticola, generalmente associate a muscoli intrinseci o estrinseci.
Caratteri della cuticola
La maggior parte delle larve dei Ditteri vive in ambiente acquatico o in microambienti rappresentati da substrati umidi di varia natura oppure dal corpo di altri organismi viventi, animali o vegetali. In rapporto alla generalità dell'ordine, sono poche le larve che vivono in ambienti secchi, come ad esempio il suolo asciutto, o direttamente all'aperto, esposte all'aria. Conseguenza di questo aspetto è che in genere le larve dei Ditteri non necessitano di particolari adattamenti che le proteggano da condizioni particolarmente sfavorevoli, come l'azione abrasiva da parte del substrato, la disidratazione, la predazione. Nella maggior parte delle larve dei Ditteri, pertanto, il tegumento è liscio e più o meno glabro, sottile e molle, poco o per nulla sclerificato, depigmentato o traslucido (Teskey, 1981, Courtney et al., 2000).
Sclerificazione
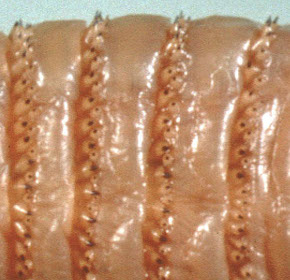
L'ispessimento e la sclerificazione del tegumento, in genere limitata a determinate aree del corpo, ricorre solo in pochi gruppi e in generale è associata a particolari abitudini e condizioni ambientali. I casi più evidenti coinvolgono le larve degli Asilidae, degli Stratiomyidae e degli Xylomyidae. Nella famiglia degli Asilidae, le larve vivono in suoli secchi e hanno un'attività predatoria. La mobilità di queste larve e le condizioni ambientali le esporrebbero alla disidratazione e all'azione abrasiva delle particelle del terreno, perciò hanno una cuticola resistente di consistenza cuoiosa (Teskey, 1981). Le larve degli Xylomyidae e di alcune sottofamiglie degli Stratiomyidae sono terrestri e vivono in materiali vari in decomposizione, come le lettiere di foglie e i residui di cortecce, oppure negli strati superficiali del suolo. Per questo motivo, probabilmente, hanno una cuticola spessa e robusta e, peculiarità di queste famiglie, ruvida per la presenza di concrezioni di carbonato di calcio (Teskey, 1981; Courtney et al., 2000; Tremblay, 2005).
In altri gruppi, la sclerificazione del tegumento non è estesa a tutto il corpo ma è circoscritta a determinate regioni. Ad esempio, le larve degli Xylophagidae hanno un tegumento leggermente chitinizzato con placche sclerificate nelle regioni dorsali e laterali del torace e nella parte dorsale dell'ultimo segmento (Teskey, 1981; Courtney et al., 2000). Le larve degli Psychodidae della sottofamiglia Psychodinae hanno in genere due placche sclerificate sulla parte dorsale di ciascun segmento toracico e del primo urite e tre per ciascuno dei segmenti addominali successivi, dal secondo al settimo (Teskey, 1981, Quate & Vockeroth, 1981 e Wagner, 1997). Le larve di molti Cecidomyiidae hanno nella parte ventrale del segmento protoracico una piccola sclerificazione variamente conformata, chiamata spatola sternale (Teskey, 1981; Gagné, 1981; Tremblay, 1991; Skuhravá, 1997). Sclerificazioni più o meno marcate si possono riscontrare per diverse famiglie di Nematoceri in strutture dell'ultimo segmento associate all'ano o agli spiracoli posteriori (Teskey, 1981). Fra le larve dei Cyclorrhapha, invece, le sclerificazioni del tegumento sono poco frequenti e limitate solo a poche famiglie (Courtney et al., 2000).
Pigmentazione
Nei Ditteri, la pigmentazione del tegumento delle larve è un carattere morfologico di limitata importanza, essendo la cuticola depigmentata, con una colorazione pallida variabile dal bianco al giallognolo, soprattutto per le forme che non vivono all'aperto (Teskey, 1981).
Pigmentazioni vere e proprie sono dovute presumibilmente alla presenza di granuli di pigmento in cromatociti del tessuto adiposo o dell'epidermide e si conoscono solo in poche famiglie di Nematoceri (Teskey, 1981; Courtney et al., 2000). In altri casi la pigmentazione si deve alla colorazione dell'emolinfa contenente carotenoidi, emocianina o emoglobina. In funzione di questi pigmenti ematici, la colorazione può variare, secondo i gruppi, dal grigio al bruno al verde, raramente al rosso (Teskey, 1981; Courtney et al., 2000).
Eventuali pigmentazioni indirette sono invece generate dalla pubescenza della cuticola e può avere varie configurazioni, talvolta con qualche utilità ai fini della determinazione tassonomica (Teskey, 1981; Courtney et al., 2000).
Autore: Alan R. Walker (Home page)
Modificata dalla foto originale su Wikimedia Commons
{kind=link}
(Licenza: Creative Commons BY-SA)
Escrescenze
Quando sono presenti, gli annessi cuticolari sono rappresentati da escrescenze di vario tipo come tubercoli, spicole, spine, peli, setole e, infine, processi molli. L'aspetto di queste formazioni cambia secondo i gruppi e può costituire un utile elemento di determinazione tassonomica. In generale sono più frequenti nelle larve dei Ditteri inferiori (Nematoceri, Brachiceri Ortorrafi e Ciclorrafi del gruppo Aschiza), mentre nei Ciclorrafi Schizofori sono generalmente circoscritte all'ultimo segmento addominale (Teskey, 1981).
Nella configurazione più semplice, gli annessi cuticolari sono rappresentati da microtrichi o da spicole di dimensioni omogenee che rivestono uniformemente tutto il corpo o estese parti di esso, carattere che si riscontra nelle larve di diverse famiglie sia tra i Nematoceri sia tra i Brachiceri (Teskey, 1981). Nelle larve di alcune famiglie si possono avere formazioni di sviluppo diverso arrangiate secondo disegni che possono essere caratteristici. Ad esempio, nei Ptychopteridae si ha una pubescenza uniforme costituita da microtrichi a cui si aggiungono peli più lunghi raggruppati in serie trasversali (Teskey, 1981). Le larve di alcuni Scatopsidae hanno bande trasversali di setole nella parte posteriore di ciascun segmento, precedute da brevi file longitudinali (Teskey, 1981; Cook, 1981).
Le larve zoofaghe di schizofori parassiti di vertebrati possono presentare formazioni cuticolari usate per ancorarsi o avanzare tra i tessuti dell'ospite: nella generalità degli Oestridae sensu lato, le larve hanno robuste spine o squamette raggruppate lungo i margini anteriori dei segmenti (Teskey, 1981; Wood, 1987; Tremblay, 1997; Minář, 2000b, 2000c, 2000d) e il loro aspetto e la loro disposizione sono utili ai fini diagnostici. Arrangiamenti simili si osservano anche in molte larve dei Calliphoridae, in genere con serie multiple di spine reclinate, disposte lungo i margini anteriori di ciascun segmento (Shewell, 1987; Tremblay, 1997; Rognes, 1998).

Sono visibili le goccioline d'acqua raccolte sulle setole secretive dorsali.
Autore: Cristophe Quintin
(Home page su Flickr)
Modificata dalla foto originale
(Licenza: Creative Commons BY-NC 2.0)
Annessi tricoidei più evidenti sono le setole, di cui variano, nell'ambito dell'ordine, la distribuzione, la dimensione, la forma (Teskey, 1981). Per quanto concerne la distribuzione, possono essere più o meno raggruppate secondo schemi di una certa regolarità oppure sparse su tutto il corpo, presenti su tutti i segmenti oppure solo in alcuni di essi. In genere hanno l'aspetto di peli più o meno sviluppati, ma possono ricorrere casi di setole con singolari conformazioni o altre caratteristiche. Ad esempio, le larve di Lutzomyia vexator (Coquillett) (Nematocera: Psychodidae) hanno caratteristiche setole a forma di clava (Quate & Vockeroth, 1981). Le larve dei Culicidae possono avere setole palmate, come negli Anophelinae, oppure ramificate, come in alcuni Culicinae (Stone, 1981; Minář, 2000a).
Le larve dei Forcipomyiinae (Nematocera: Ceratopogonidae), sottofamiglia comprendente i generi Forcipomyia Meigen e Atrichopogon Kieffer, hanno un complesso sistema di setole di diversa conformazione presenti su tutti i segmenti toracici e addominali. In generale, le setole sono distribuite su più serie longitudinali, di cui quattro dorsali inserite su tubercoli più o meno pronunciati (Downes & Wirth, 1981; Boorman, 1997). Le setole delle due serie centrali mediane risultano essere associate a meccanorecettori (Urbanek et al., 2015), ma nel genere Forcipomyia, la cellula tricogena ha acquisito una funzione ghiandolare e secerne, attraverso l'apice spatolato della setola, una sostanza igroscopica (Urbanek et al., 2011). Conseguenza di questa peculiarità anatomo-fisiologica è che sul dorso sono spesso presenti goccioline d'acqua all'apice di queste setole che rendono singolare l'aspetto di queste larve (Murray, All about Forcipomyiinae, galleria fotografica). La principale funzione di queste secrezioni ghiandolari sarebbe quella di preservare le larve dall'essiccazione e al tempo stesso facilitare la respirazione cutanea: le gocce d'acqua scendono lungo la setola e si espandono sulla cuticola della larva ricoprendola con un velo umido (Urbanek et al., 2015). In effetti, questa caratteristica è riscontrata nelle larve terrestri delle specie di questo genere, frequenti sotto le cortecce degli alberi e tra i muschi (Boorman, 1997).
Bibliografia
- Boorman, J. (1997) 2.21. Family Ceratopogonidae: 349-368. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 2. Nematocera and Lower Brachycera, Op. cit.
- Cook, E.F. (1981) Scatopsidae: 313-319. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera, Volume 1, Op. cit.
- Courtney, W.G.; Sinclair, B.J. & Meier, R. (2000) 1.4. Morphology and terminology of Diptera larvae: 85-163. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 1. General and Applied Dipterology, Op. cit.
- Downes, J.A. & Wirth, W.W. (1981) Ceratopogonidae: 393-421. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera, Volume 1, Op. cit.
- Gagné, R.J. (1981) Cecidomyiidae: 257-292. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera, Volume 1, Op. cit.
- Minář, J. (2000a) A.5. Family Culicidae: 93-111. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 4. Appendix, Op. cit.
- Minář, J. (2000b) A.14. Family Gasterophylidae: 455-466. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 4. Appendix, Op. cit.
- Minář, J. (2000c) A.15. Family Oestridae: 467-478. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 4. Appendix, Op. cit.
- Minář, J. (2000d) A.16. Family Hypodermatidae: 479-494. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 4. Appendix, Op. cit.
- Quate, L.W. & Vockeroth, J.R. (1981) Psychodidae: 293-300. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera, Volume 1, Op. cit.
- Rognes, K. (1998) 3.51. Family Calliphoridae: 617-648. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 3. Higher Brachycera, Op. cit.
- Shewell, G.E. (1987) Calliphoridae: 1133-1145. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera, Volume 2, Op. cit.
- Skuhravá, M. (1997) 2.7. Family Cecidomyiidae: 71-204. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 2. Nematocera and Lower Brachycera, Op. cit.
- Stone, A. (1981) Culicidae: 341-350. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera, Volume 1, Op. cit.
- Teskey, H.J. (1981) Morphology and terminology - Larvae: 65-88. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera, Volume 1, Op. cit.
- Tremblay, E. (1991) Entomologia applicata. Volume III Parte prima, Op. cit.
- Tremblay, E. (1997) Entomologia applicata. Volume III Parte terza, Ditteri Brachiceri (Caliptrati), Sifonatteri e Strepsitteri, Op. cit.
- Tremblay, E. (2005) Entomologia applicata. Volume III Parte seconda, Ditteri Brachiceri (Caliptrati esclusi), Op. cit.
- Urbanek, A.; Richert, M.; Giłka, W. & Szadziewski, R. (2015) Morphology and histology of secretory setae in terrestrial larvae of biting midges of the genus Forcipomyia (Diptera: Ceratopogonidae). Arthropod Structure & Development 40 (6): 485-494.
- Urbanek, A.; Szadziewski, R. & Giłka, W. (2015) Ultrastructure of mediodorsal setae in biting midge larvae of the genus Atrichopogon Kieffer with notes on their biological significance (Diptera: Ceratopogonidae). Arthropod Structure & Development 44 (1): 10-20.
- Wagner, R. (1997) 2.8. Family Psychodidae: 205-226. In Papp, L. & Darvas, B. (a cura di) Contributions to a Manual of Palaearctic Diptera. Volume 2. Nematocera and Lower Brachycera, Op. cit.
- Wood, D.M. (1987) Oestridae: 1147-1158. In McAlpine, J.F.; Peterson, B.V.; Shewell, G.E; Teskey, H.J.; Vockeroth, J.R. & Wood, D.M. (a cura di) Manual of Nearctic Diptera, Volume 2, Op. cit.
Risorse Web
- Andy Murray: All about Forcipomyiinae. In A Caos of Delight. The wonderful world of soil mesofauna. Ultimo accesso: 28 maggio 2019.

Licenza Creative Commons BY-NC-SA
(BY: Attribuzione - NC: Non commerciale - SA: Condividi allo stesso modo).